Saturday, 2 June 2018
On Babylon the great:The Watchtower Society's commentary.
BABYLON THE GREAT:
Among John’s visions recorded in the book of Revelation appear pronouncements of judgment against “Babylon the Great,” as well as a description of her and of her downfall.—Re 14:8; 16:19; chaps 17, 18; 19:1-3.
In Revelation 17:3-5, Babylon the Great is described as a woman arrayed in purple and scarlet, richly adorned, and sitting upon a scarlet-colored wild beast having seven heads and ten horns. Upon her forehead a name is written, “a mystery: ‘Babylon the Great, the mother of the harlots and of the disgusting things of the earth.’” She is also depicted as sitting on “many waters” representing “peoples and crowds and nations and tongues.”—Re 17:1-15.
The luxury and the dominion attributed to Babylon the Great do not allow for simply equating her with the literal city of Babylon in Mesopotamia. After ancient Babylon fell to Cyrus the Persian in 539 B.C.E., it lost its position as a dominant world power, its captives, including the Jews, being freed. Although the city continued to exist even beyond the days of the apostles, and hence existed in John’s day, it was no longer a city of world importance, and it eventually fell into decay and utter ruin. Thus, Babylon the Great must be viewed as a symbolic city, one of which the literal city of Babylon was the prototype. Because the ancient city gives the mystic city its name, it is helpful to consider briefly the outstanding features of Babylon on the Euphrates, features that provide clues as to the identity of the symbolic city of John’s vision.
Characteristics of Ancient Babylon. The founding of the city of Babylon on the Plains of Shinar was concurrent with the attempt at building the Tower of Babel. (Ge 11:2-9) The popular cause to be advanced by the tower and city construction was, not the exaltation of God’s name, but that the builders might “make a celebrated name” for themselves. The ziggurat towers uncovered not only in the ruins of ancient Babylon but elsewhere in Mesopotamia would seem to confirm the essentially religious nature of the original tower, whatever its form or style. The decisive action taken by Jehovah God to overthrow the temple construction clearly condemns it as of a false religious origin. Whereas the Hebrew name given the city, Babel, means “Confusion,” the Sumerian name (Ka-dingir-ra) and the Akkadian name (Bab-ilu) both mean “Gate of God.” Thus the remaining inhabitants of the city altered the form of its name to avoid the original condemnatory sense, but the new or substitute form still identified the city with religion.
The Bible lists Babel first when giving the ‘beginning of Nimrod’s kingdom.’ (Ge 10:8-10) Throughout the Hebrew Scriptures the ancient city of Babylon is featured prominently as the longtime enemy of Jehovah God and his people.
Though Babylon became the capital of a political empire in the seventh and sixth centuries B.C.E., it was outstandingly prominent during its entire history as a religious center from which religious influence radiated in many directions.
Professor Morris Jastrow, Jr., in his work The Religion of Babylonia and Assyria (1898, pp. 699-701), says regarding this: “In the ancient world, prior to the rise of Christianity, Egypt, Persia, and Greece felt the influence of the Babylonian religion. . . . In Persia, the Mithra cult reveals the unmistakable influence of Babylonian conceptions; and if it be recalled what a degree of importance the mysteries connected with this cult acquired among the Romans, another link will be added connecting the ramifications of ancient culture with the civilization of the Euphrates Valley.” In conclusion he refers to “the profound impression made upon the ancient world by the remarkable manifestations of religious thought in Babylonia and by the religious activity that prevailed in that region.”
Babylon’s religious influence is traced eastward to India in the book New Light on the Most Ancient East, by archaeologist V. Childe (1957, p. 185). Among other points he states: “The swastika and the cross, common on stamps and plaques, were religious or magical symbols as in Babylonia and Elam in the earliest prehistoric period, but preserve that character also in modern India as elsewhere.” Thus, ancient Babylon’s religious influence spread out to many peoples and nations, much farther and with greater potency and endurance than did her political strength.
Like mystic Babylon, the ancient city of Babylon, in effect, sat on the waters, located, as it was, astride the Euphrates River and having various canals and water-filled moats. (Jer 51:1, 13; Re 17:1, 15) These waters served as a defense to the city, and they provided the thoroughfares upon which ships brought wealth and luxuries from many sources. Notably, the water of the Euphrates is depicted as drying up prior to Babylon the Great’s experiencing the wrath of divine judgment.—Re 16:12, 19.
Distinguishing Features of Mystic Babylon. The symbolic woman bearing the name Babylon the Great is “the great city that has a kingdom over the kings of the earth,” a kingdom that allows her, in effect, to sit on “peoples and crowds and nations and tongues.” (Re 17:1, 15, 18) A kingdom over other kingdoms and nations is what is defined as an “empire.” Babylon the Great places herself above earthly kings, exercising power and influence over them. She rides the symbolic seven-headed beast, beasts being used elsewhere in the Bible as symbols of political world powers.—See BEASTS, SYMBOLIC.
Some scholars assume that Babylon the Great is a political empire, either Babylon or Rome. We have already seen that Babylon as a political empire had long since ceased to exist when John received his prophetic vision. As to Rome, the nature of its political rule does not harmonize with the description of Babylon the Great’s course and her methods of dominating. She is a harlot, committing fornication with the kings of the earth, making them drunk with the wine of her fornication, misleading the nations by her “spiritistic practice.” (Re 17:1, 2; 18:3, 23) Rome’s dominion, by contrast, was gained and maintained by its ironlike military might and its firm application of Roman law among its provinces and colonies. Recognizing this fact, The Interpreter’s Dictionary of the Bible says: “It is not sufficient to identify Rome and Babylon. Babylon embraces more than one empire or culture. It is defined rather by dominant idolatries than by geographical or temporal boundaries. Babylon is coextensive with the kingdom of that beast which has corrupted and enslaved mankind, and whom the Lamb must conquer (Rev. 17:14) if mankind is to be freed.”—Edited by G. Buttrick, 1962, Vol. 1, p. 338.
The symbol of a harlot or a fornicatrix is used frequently in the Hebrew Scriptures. The nation of Israel was warned against entering into covenant relations with the nations of Canaan because this would lead them to commit “immoral intercourse [“play the harlot,” RS] with their gods.” (Ex 34:12-16) Both Israel and Judah apostatized from the true worship of Jehovah God and were condemned by him as having engaged in harlotry, prostituting themselves to the political nations and their gods. (Isa 1:21; Jer 3:6-10, 13; Eze 16:15-17, 28, 29, 38; Ho 6:10; 7:11; 8:9, 10) It may be noted here that God was not viewing Israel or Judah as mere political entities entering into relations with other political governments. Instead God reprimanded them on the basis of their being in a sacred covenant with him, hence responsible to be a holy people devoted to him and his pure worship.—Jer 2:1-3, 17-21.
A similar usage of this figure is found in the Christian Greek Scriptures. The Christian congregation is likened to a virgin espoused to Christ as her Head and King. (2Co 11:2; Eph 5:22-27) The disciple James warned Christians against committing spiritual adultery through friendship with the world. (Jas 4:4; compare Joh 15:19.) The fornications of Babylon the Great and her “daughters” are of a similar nature and not some unique exception. (The term “daughters” at times is employed in the Bible to refer to the suburbs or surrounding towns of a city or metropolis, as the “dependent towns” [literally, “daughters” in Hebrew] of Samaria and Sodom; see Eze 16:46-48.)
An additional significant factor is that when Babylon the Great goes down under the devastating attack of the ten horns of the symbolic wild beast, her fall is mourned by her companions in fornication, the kings of the earth, and also by the merchants and shippers who dealt with her in supplying luxurious commodities and gorgeous fineries. While these political and commercial representatives survive her desolation, notably no religious representatives are depicted as still on the scene to share in mourning her downfall. (Re 17:16, 17; 18:9-19) The kings of the earth are shown as having judgment executed upon them sometime after mystic Babylon’s annihilation, and their destruction comes, not from the “ten horns,” but from the sword of the King of kings, the Word of God.—Re 19:1, 2, 11-18.
A further distinguishing characteristic of Babylon the Great is her drunkenness, she being pictured as “drunk with the blood of the holy ones and with the blood of the witnesses of Jesus.” (Re 17:4, 6; 18:24; 19:1, 2) She thus is the spiritual counterpart of the ancient city of Babylon, expressing the same enmity toward the true people of God. Significantly, it was to the charge of religious leaders that Jesus laid the responsibility for “all the righteous blood spilled on earth, from the blood of righteous Abel to the blood of Zechariah.” While those words were addressed to religious leaders from among Jesus’ own race, the Jewish nation, and while persecution against Jesus’ followers was particularly intense from that sector for a time, history shows that thereafter the opposition to genuine Christianity came from other sources (the Jews themselves suffering considerable persecution).—Mt 23:29-35.
All the above factors are significant, and they must all be considered in arriving at a true picture of symbolic Babylon the Great and what it represents.
Among John’s visions recorded in the book of Revelation appear pronouncements of judgment against “Babylon the Great,” as well as a description of her and of her downfall.—Re 14:8; 16:19; chaps 17, 18; 19:1-3.
In Revelation 17:3-5, Babylon the Great is described as a woman arrayed in purple and scarlet, richly adorned, and sitting upon a scarlet-colored wild beast having seven heads and ten horns. Upon her forehead a name is written, “a mystery: ‘Babylon the Great, the mother of the harlots and of the disgusting things of the earth.’” She is also depicted as sitting on “many waters” representing “peoples and crowds and nations and tongues.”—Re 17:1-15.
The luxury and the dominion attributed to Babylon the Great do not allow for simply equating her with the literal city of Babylon in Mesopotamia. After ancient Babylon fell to Cyrus the Persian in 539 B.C.E., it lost its position as a dominant world power, its captives, including the Jews, being freed. Although the city continued to exist even beyond the days of the apostles, and hence existed in John’s day, it was no longer a city of world importance, and it eventually fell into decay and utter ruin. Thus, Babylon the Great must be viewed as a symbolic city, one of which the literal city of Babylon was the prototype. Because the ancient city gives the mystic city its name, it is helpful to consider briefly the outstanding features of Babylon on the Euphrates, features that provide clues as to the identity of the symbolic city of John’s vision.
Characteristics of Ancient Babylon. The founding of the city of Babylon on the Plains of Shinar was concurrent with the attempt at building the Tower of Babel. (Ge 11:2-9) The popular cause to be advanced by the tower and city construction was, not the exaltation of God’s name, but that the builders might “make a celebrated name” for themselves. The ziggurat towers uncovered not only in the ruins of ancient Babylon but elsewhere in Mesopotamia would seem to confirm the essentially religious nature of the original tower, whatever its form or style. The decisive action taken by Jehovah God to overthrow the temple construction clearly condemns it as of a false religious origin. Whereas the Hebrew name given the city, Babel, means “Confusion,” the Sumerian name (Ka-dingir-ra) and the Akkadian name (Bab-ilu) both mean “Gate of God.” Thus the remaining inhabitants of the city altered the form of its name to avoid the original condemnatory sense, but the new or substitute form still identified the city with religion.
The Bible lists Babel first when giving the ‘beginning of Nimrod’s kingdom.’ (Ge 10:8-10) Throughout the Hebrew Scriptures the ancient city of Babylon is featured prominently as the longtime enemy of Jehovah God and his people.
Though Babylon became the capital of a political empire in the seventh and sixth centuries B.C.E., it was outstandingly prominent during its entire history as a religious center from which religious influence radiated in many directions.
Professor Morris Jastrow, Jr., in his work The Religion of Babylonia and Assyria (1898, pp. 699-701), says regarding this: “In the ancient world, prior to the rise of Christianity, Egypt, Persia, and Greece felt the influence of the Babylonian religion. . . . In Persia, the Mithra cult reveals the unmistakable influence of Babylonian conceptions; and if it be recalled what a degree of importance the mysteries connected with this cult acquired among the Romans, another link will be added connecting the ramifications of ancient culture with the civilization of the Euphrates Valley.” In conclusion he refers to “the profound impression made upon the ancient world by the remarkable manifestations of religious thought in Babylonia and by the religious activity that prevailed in that region.”
Babylon’s religious influence is traced eastward to India in the book New Light on the Most Ancient East, by archaeologist V. Childe (1957, p. 185). Among other points he states: “The swastika and the cross, common on stamps and plaques, were religious or magical symbols as in Babylonia and Elam in the earliest prehistoric period, but preserve that character also in modern India as elsewhere.” Thus, ancient Babylon’s religious influence spread out to many peoples and nations, much farther and with greater potency and endurance than did her political strength.
Like mystic Babylon, the ancient city of Babylon, in effect, sat on the waters, located, as it was, astride the Euphrates River and having various canals and water-filled moats. (Jer 51:1, 13; Re 17:1, 15) These waters served as a defense to the city, and they provided the thoroughfares upon which ships brought wealth and luxuries from many sources. Notably, the water of the Euphrates is depicted as drying up prior to Babylon the Great’s experiencing the wrath of divine judgment.—Re 16:12, 19.
Distinguishing Features of Mystic Babylon. The symbolic woman bearing the name Babylon the Great is “the great city that has a kingdom over the kings of the earth,” a kingdom that allows her, in effect, to sit on “peoples and crowds and nations and tongues.” (Re 17:1, 15, 18) A kingdom over other kingdoms and nations is what is defined as an “empire.” Babylon the Great places herself above earthly kings, exercising power and influence over them. She rides the symbolic seven-headed beast, beasts being used elsewhere in the Bible as symbols of political world powers.—See BEASTS, SYMBOLIC.
Some scholars assume that Babylon the Great is a political empire, either Babylon or Rome. We have already seen that Babylon as a political empire had long since ceased to exist when John received his prophetic vision. As to Rome, the nature of its political rule does not harmonize with the description of Babylon the Great’s course and her methods of dominating. She is a harlot, committing fornication with the kings of the earth, making them drunk with the wine of her fornication, misleading the nations by her “spiritistic practice.” (Re 17:1, 2; 18:3, 23) Rome’s dominion, by contrast, was gained and maintained by its ironlike military might and its firm application of Roman law among its provinces and colonies. Recognizing this fact, The Interpreter’s Dictionary of the Bible says: “It is not sufficient to identify Rome and Babylon. Babylon embraces more than one empire or culture. It is defined rather by dominant idolatries than by geographical or temporal boundaries. Babylon is coextensive with the kingdom of that beast which has corrupted and enslaved mankind, and whom the Lamb must conquer (Rev. 17:14) if mankind is to be freed.”—Edited by G. Buttrick, 1962, Vol. 1, p. 338.
The symbol of a harlot or a fornicatrix is used frequently in the Hebrew Scriptures. The nation of Israel was warned against entering into covenant relations with the nations of Canaan because this would lead them to commit “immoral intercourse [“play the harlot,” RS] with their gods.” (Ex 34:12-16) Both Israel and Judah apostatized from the true worship of Jehovah God and were condemned by him as having engaged in harlotry, prostituting themselves to the political nations and their gods. (Isa 1:21; Jer 3:6-10, 13; Eze 16:15-17, 28, 29, 38; Ho 6:10; 7:11; 8:9, 10) It may be noted here that God was not viewing Israel or Judah as mere political entities entering into relations with other political governments. Instead God reprimanded them on the basis of their being in a sacred covenant with him, hence responsible to be a holy people devoted to him and his pure worship.—Jer 2:1-3, 17-21.
A similar usage of this figure is found in the Christian Greek Scriptures. The Christian congregation is likened to a virgin espoused to Christ as her Head and King. (2Co 11:2; Eph 5:22-27) The disciple James warned Christians against committing spiritual adultery through friendship with the world. (Jas 4:4; compare Joh 15:19.) The fornications of Babylon the Great and her “daughters” are of a similar nature and not some unique exception. (The term “daughters” at times is employed in the Bible to refer to the suburbs or surrounding towns of a city or metropolis, as the “dependent towns” [literally, “daughters” in Hebrew] of Samaria and Sodom; see Eze 16:46-48.)
An additional significant factor is that when Babylon the Great goes down under the devastating attack of the ten horns of the symbolic wild beast, her fall is mourned by her companions in fornication, the kings of the earth, and also by the merchants and shippers who dealt with her in supplying luxurious commodities and gorgeous fineries. While these political and commercial representatives survive her desolation, notably no religious representatives are depicted as still on the scene to share in mourning her downfall. (Re 17:16, 17; 18:9-19) The kings of the earth are shown as having judgment executed upon them sometime after mystic Babylon’s annihilation, and their destruction comes, not from the “ten horns,” but from the sword of the King of kings, the Word of God.—Re 19:1, 2, 11-18.
A further distinguishing characteristic of Babylon the Great is her drunkenness, she being pictured as “drunk with the blood of the holy ones and with the blood of the witnesses of Jesus.” (Re 17:4, 6; 18:24; 19:1, 2) She thus is the spiritual counterpart of the ancient city of Babylon, expressing the same enmity toward the true people of God. Significantly, it was to the charge of religious leaders that Jesus laid the responsibility for “all the righteous blood spilled on earth, from the blood of righteous Abel to the blood of Zechariah.” While those words were addressed to religious leaders from among Jesus’ own race, the Jewish nation, and while persecution against Jesus’ followers was particularly intense from that sector for a time, history shows that thereafter the opposition to genuine Christianity came from other sources (the Jews themselves suffering considerable persecution).—Mt 23:29-35.
All the above factors are significant, and they must all be considered in arriving at a true picture of symbolic Babylon the Great and what it represents.
That pesky second law continues to bedevil abiogenesis
Thermodynamics of the Origin of Life
Brian Miller
Brian Miller
Popular articles on origin-of-life research have often portrayed the field as constantly advancing and quickly converging on a purely materialistic explanation for the first autonomous cell. However, the true state of affairs is far grimmer. The highpoint of optimism was actually in the 1950s when Stanley Miller’s experiment produced several of the building blocks of life. Unfortunately, geologists later recognized that the gases used in that experiment did not match the chemistry of the early Earth. When redone with those actually present, the experiments produced few building blocks in sufficient quantities to contribute to origin-of-life scenarios. Moreover, close examination of modern attempts to synthesize life’s precursors reveals that the formation of many of them on the early Earth in significant quantities is virtually impossible. However, the more fundamental challenge of thermodynamics dwarfs even these hurdles.
Common arguments about the origin of life have traditionally focused on the unlikelihood of life forming by chance. Perhaps most famously, physicist Fred Hoyle calculated the probability of a cell coalescing to be roughly 1 part in 10 to the power of 40,000. He compared this probability to the chances of a tornado plowing through a junkyard and assembling a jet airplane. The concept of probability is closely linked to that of entropy, since probability is proportional to the number of configurations (N) in which some state could occur, and entropy is proportional to the log of N. As an example, the number of ways water molecules can arrange themselves in the solid state is much smaller than the number ways in the liquid or gas states, so ice is the state with the lowest entropy. Due to this connection, the probability argument is often restated that nature tends to move from states of lower entropy to higher entropy, which simply means that nature moves towards states which are highly probable. This tendency is known as the second law of thermodynamics.
Such arguments sound impressive at first, but they are actually incomplete. Nearly all researchers recognize that the first cell could not have come about by chance. They instead believe that some physical processes helped to beat the odds. As an analogy, one could never role one thousand sixes in a row with fair dice. However, if the dice were loaded, that outcome could be quite likely or even close to guaranteed. Analogously, some systems do, in fact, naturally move from states of higher entropy to those of lower entropy (i.e., seemingly low probability) if the lower-entropy states are highly biased to occur. Such a bias is created by a second driving tendency. Namely, nature tends to move from states of higher energy to those of lower energy. For instance, rocks roll downhill, since lower altitude corresponds to lower gravitational energy. Likewise, molecules of water attract each other, so ice is a lower energy state since the water molecules are on average closer to each other than in the liquid or gas states. At low enough temperatures, this attraction overcomes the tendency to move toward higher entropy resulting in water freezing.
However, even in these cases of locally decreasing entropy, the second law of thermodynamics is not violated, for the changes are always exothermic — heat is released. The heat leaving the local system (e.g., a cup of freezing water) and entering the surrounding environment increases the latter’s entropy by an amount greater than the entropy decrease of the local system. Therefore, the total entropy of the universe increases. The problem for all theories of origin of life now becomes quite evident. The simplest functional cell compared to its most basic building blocks has both lower entropy and higher energy. And, natural systems never both decrease in entropy and increase in energy at the same time. Such an event would be like rolling countless sixes in a row when the dice are strongly loaded against rolling even one. Therefore, the origin of life through purely natural processes would seem as implausible as water on a hot summer day spontaneously freezing or a river flowing unaided uphill for thousands of miles.
Physicists and chemists often combine entropy and energy (or enthalpy) together into what is called the free energy of a system. The change of free energy is always negative for spontaneous changes (e.g., wood burning or ice melting in summer), and it directly relates to the total increase in entropy of the universe. The challenge for the origin of life is then explaining how billions of atoms could spontaneously come together into a state of significantly higher free energy. The chance of such an event happening through thermal fluctuations was calculated by biophysicist Harold Morowitz to be less than 1 part in 10 to the power of a hundred million This number comes directly from an estimation of the free energy of life, and the free energy is a path-independent function. Therefore, this maximum probability estimate is not meaningfully dependent on the theoretical route that led to life (e.g., protein-first versus RNA world) or on the number of steps involved. The probability is always essentially zero.
At face value this thermodynamic analysis for the origin of life would seem to negate any possible materialistic solution to the problem. However, theorists have long recognized one remaining loophole. Morowitz’s calculations assumed that the system was in a state near equilibrium. For instance, some lightning bolt could have struck a pond of prebiotic chemicals at night causing atoms to come together into different configurations. The pond would then quickly settle back into a calm state where the temperature, concentrations, and other variables would remain fairly uniform. However, many argue that the origin of life took place in a system strongly driven away from equilibrium, such as a pond subjected to intense sunlight or the bottom of the ocean near a hydrothermal vent flooding its surroundings with superheated water and high-energy chemicals. Such settings are commonly referred to as non-equilibrium dissipative systems. Their common characteristic is that classical thermodynamics breaks down, so the previous analyses do not completely hold. Instead, principles of non-equilibrium thermodynamics must be applied, which are far more complex and less well understood. Moreover, the energy from these outside sources is hoped to enable the free-energy barrier to be overcome.
However, such appeals to non-equilibrium systems do little to solve the basic thermodynamic problems. First, no system could be maintained far from equilibrium for more than a limited amount of time. The sun is only out during the day, and superheated water at the bottom of the ocean would eventually migrate away from any hydrothermal vents. Any progress made toward forming a cell would be lost as the system reverted toward equilibrium (lower free energy) and thus away from any state approaching life. Second, the input of raw solar, thermal, or other forms of energy actually increase the entropy of the system, thus moving it in the wrong direction. For instance, the ultraviolet light from the sun or heat from hydrothermal vents would less easily form the complex chemical structures needed for life than break them apart. Finally, in non-equilibrium systems the differences in temperature, concentrations, and other variables act as thermodynamic forces which drive heat transfer, diffusion, and other thermodynamic flows. These flows create microscopic sources of entropy production, again moving the system away from any reduced-entropy state associated with life. In short, the processes occurring in non-equilibrium systems, as in their near-equilibrium counterparts, generally do the opposite of what is actually needed.
It's still design all the way down.
Life Exponential: Life Exhibits Intelligent Design at Many Levels
Jonathan Wells
Jonathan Wells
A human cell contains two sets of DNA, each consisting of about three billion subunits called “nucleotides.” There are four different nucleotides, and they can be arranged in many different ways, so DNA is quite complex. Most of our DNA, however, must be arranged in a very specific way to provide the information a cell uses to make RNAs and proteins. Mathematician William Dembski has called this “complex specified information.”1
Complexity (such as we see in a pile of autumn leaves) can arise spontaneously from unguided natural processes, but complex specified information cannot. The only known source of complex specified information is an intelligent mind, which can envision a goal and arrange things to actualize that goal — in this case, a living cell. “Because mind or intelligent design is a necessary cause of an information-rich system,” philosopher of science Stephen Meyer wrote in 1998, “the specifically arranged nucleotide sequences — the encoded information — in DNA imply the past action of an intelligent mind, even if such mental agency cannot be directly observed.”2
The Need for Spatial Information
So DNA carries biological information, and that information points to design. But DNA is not the only carrier of information in a living cell. When DNA is transcribed into RNAs, most of those RNAs must be transported to specific locations in the cell before they can function properly. Some RNAs are tagged with sequences called “zipcodes” that specify the “addresses” to which they are to be transported. Like the zipcode on a letter you put in a mailbox, however, an RNA zipcode is meaningless unless it corresponds to a pre-existing address. Like the geographical addresses in a postal system, the cellular destinations of RNAs and proteins must be specified independently of their zipcodes, before they are “mailed.”
Many of those destinations are specified by molecules embedded in cell membranes. Scientists originally thought that proteins could diffuse freely in a membrane, like boats floating on the sea. It is now known, however, that many membrane proteins are arranged in non-random patterns that can be quite stable. Such patterns provide the cell with spatial information that goes beyond the information in DNA.
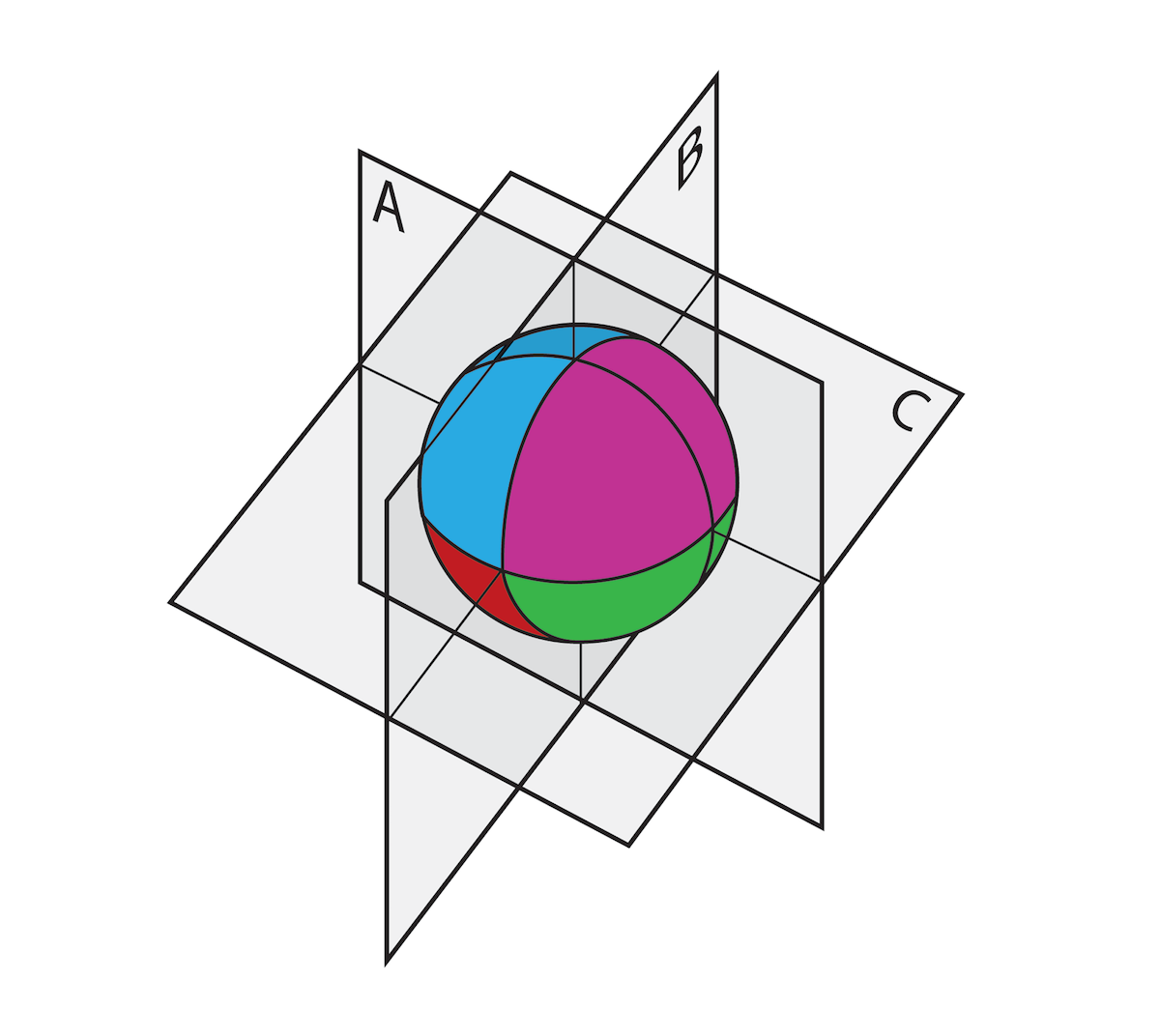
After fertilization, a frog egg is invisibly divided into regions distinguished by (among other things) spatially localized RNAs just inside the cell membrane in a zone called the “cortex.” In the accompanying drawing, four regions are indicated by different colors. After the nucleus duplicates, the fertilized egg divides into two daughter cells. If the plane of division corresponds to A, each daughter cell inherits not only a nucleus but also portions of all four regions of cortical information. If those two cells are then separated, each can develop into a complete frog. But if the plane of division corresponds to B or C, the daughter cells do not inherit a full set of cortical information, and their development is blocked even if they each contain all the necessary DNA.
Illustration: Regions of spatial information in a fertilized frog egg.
Other Carriers of Spatial Information
Regional differences in cells and embryos can be specified in other ways besides localization of RNAs in the cortex. Two of those ways have been studied in great detail: the “sugar code” and the “bioelectric code.”
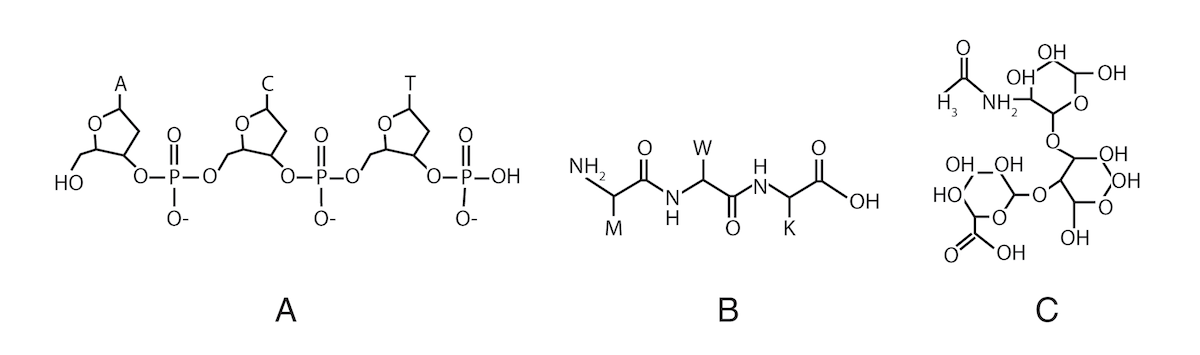
Most proteins in living cells — including those in membranes — are chemically bonded to carbohydrates called “glycans” (from the Greek word for “sweet”). The nucleotides in DNA are linked together end-to-end in a linear molecule, so DNA sequence information is one-dimensional. In living cells, the subunits in proteins (with a few exceptions) are also linked in a linear chain. But glycans can be linked together in complex three-dimensional ways, so their information-carrying capacity exceeds that of DNA and proteins by many orders of magnitude.3
Illustration: Three subunits linked together in DNA (A), a protein (B) and a glycan (C).
The information carried by glycans has been called the “sugar code.”4 The sugar code is “interpreted” by proteins called lectins, which “recognize” specific three-dimensional structures of glycan molecules. Glycans and lectins play an essential role in communication among cells and help to guide cell movements in a developing embryo. Experiments have shown that membrane patterns of glycans change in the course of embryo development.5
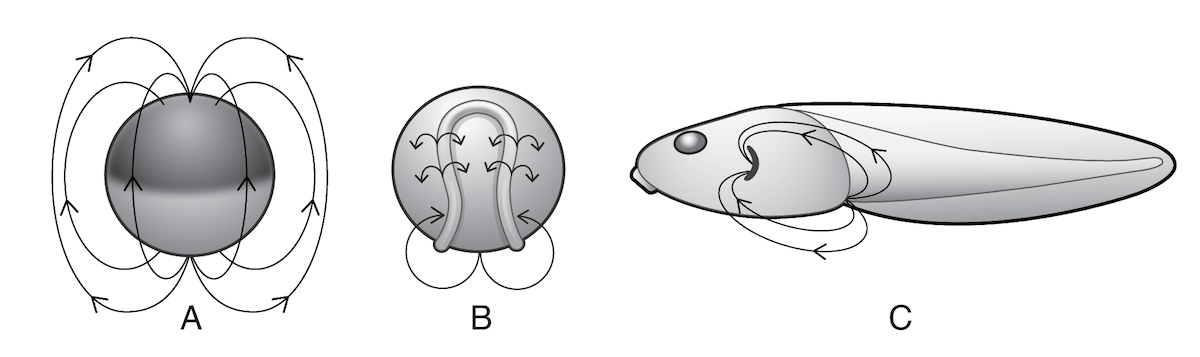
In addition to the sugar code, probably all living cells (not just nerve and muscle cells) generate electric fields across their membranes. They do this by pumping charged ions through channels in their membranes, creating a “bioelectric field.” The pattern of membrane channels determines the form of the bioelectric field, and the form of the field changes during embryo development.
Illustration: Some of the bioelectric fields (shown by arrows) in a developing frog embryo.
Bioelectric fields are correlated with important developmental events. In frog embryos, for example, large ionic currents start flowing out of the sites where the hind limbs will develop long before the limbs actually appear.
Many experiments conducted since the 1980s have confirmed that disrupting bioelectric fields causes disruptions in development. For example, frog embryos normally generate an electric field in the head-to-tail direction. If an artificial field of the same magnitude is applied in the opposite direction, or if ion channels that generate the field are blocked, the result is abnormal head and eye development. The spots where eyes normally form are more highly charged than the surrounding tissue; if the charge is neutralized, the eyes that develop are small or deformed. Sometimes, eyes develop elsewhere on the tadpole’s body, including its tail.5
How do electric fields influence development? In the 1980s, biologists exposed embryonic cells to artificial electric fields of the same strength as ones the cells generate naturally. Some types of cells migrated toward the positive pole, while other types migrated to the negative pole, suggesting that one way bioelectric fields affect embryo development is by directing cell movements.
In 1995, biologists Riyi Shi and Richards Borgens concluded that bioelectric fields “may provide a three dimensional coordinate system” that helps to specify form in embryos.6 In 2013, biologists AiSun Tseng and Michael Levin wrote that such fields may provide “templates of shape,” and that a full understanding of embryo development will probably require cracking the “bioelectric code.”7
The Membrane Code
So localized RNAs in the cortex, glycan patterns on the membrane, and bioelectric fields generated by ion channels in the membrane all carry spatial information. Although individual molecules may be specified by DNA, their three-dimensional patterns are not. Taken together, these patterns constitute a “membrane code” that is independent of DNA sequences.
In 1983, biologist Robert Poyton suggested that biological membranes carry “spatial memory,” the units of which are spatially localized proteins. Poyton wrote: “Realizing that genetic memory is one-dimensional, along a DNA molecule, whereas spatial memory is likely to be two-dimensional, along membrane surfaces, and three-dimensional within the cellular interior, it is probable that spatial memory is more complicated and diverse than genetic memory.”8
In 2004, biologist Thomas Cavalier-Smith wrote that the idea that DNA contains all the information needed to make an organism “is simply false.” According to Cavalier-Smith, membranes provide “chemically specific two-dimensional surfaces with mutually conserved topological relationships in the three spatial dimensions” that play “a key role in the mechanisms that convert the linear information of DNA into the three-dimensional shapes of single cells and multicellular organisms. Animal development creates a complex three-dimensional multicellular organism not by starting from the linear information in DNA… but always starting from an already highly complex three-dimensional unicellular organism, the fertilized egg.”9
So the membrane code carries essential biological information that is independent of DNA sequence information. Yet we often hear that embryo development is directed by a program in DNA. Why?
Beyond DNA
James Watson and Francis Crick’s Nobel Prize-winning discovery of the molecular structure of DNA in 1953 seemed to provide a molecular basis not only for heredity but also for embryo development. Cells replicate their DNA before they divide and (usually) pass on a complete set of their DNA sequences to each of their descendants. Cells then use DNA sequences as templates for the transcription of RNAs, some of which are then translated into proteins.
In the mid 20th century, biology was dominated by neo-Darwinism, a system of thought that combined evolution and genetics and attributed new variations to genetic mutations. An underlying assumption of neo-Darwinism is that evolution and development are due entirely to unguided material processes. After 1953, this materialistic assumption led to the view that “DNA makes RNA makes protein makes us,” which has been called the Central Dogma of molecular biology.
In 1970, molecular biologist (and materialist) Jacques Monod said that with the Central Dogma, “and the understanding of the random physical basis of mutation that molecular biology has also provided, the mechanism of Darwinism is at last securely founded. And man has to understand that he is a mere accident.”9
But the existence of the membrane code shows that the Central Dogma is false. And the materialistic idea that evolution is unguided cannot account for the complex specified information in DNA, much less for the extensive complex specified information in the membrane code. Just as the information in DNA points to design, so does the information beyond DNA.
Notes:
- William A. Dembski, “Intelligent design as a theory of information,” February 20, 1997.
- Stephen C. Meyer, “DNA by design,” Rhetoric and Public Affairs 1 (1998): 519-556.
- Roger A. Laine, “A calculation of all possible oligosaccharide isomers both branched and linear yields 1.05 x 1012 structures for a reducing hexasaccharide,” Glycobiology 4 (1994): 759-767.
- Hans-Joachim Gabius, “Biological information transfer beyond the genetic code: The sugar code,” Naturwissenschaften 87 (2000): 108-121.
- Jonathan Wells, “Membrane patterns carry ontogenetic information that is specified independently of DNA,” Bio-Complexity 2014 (2): 1-28.
- Riyi Shi and Richard B. Borgens, “Three-dimensional gradients of voltage during development of the nervous system as invisible coordinates for the establishment of embryonic pattern,” Developmental Dynamics 202 (1995): 101-114.
- AiSun Tseng and Michael Levin, “Cracking the bioelectric code: Probing endogenous ionic controls of pattern formation,” Communicative and Integrative Biology 6 (2013): e22595
- Robert O. Poyton, “Memory and membranes: The expression of genetic and spatial memory during the assembly of organelle macrocompartments,” Modern Cell Biology 2 (1983): 15-72.
- Thomas Cavalier-Smith, “The membranome and membrane heredity in development and evolution,” pp. 335-351 in Robert P. Hirt and David S. Horner (editors), Organelles, Genomes and Eukaryote Phylogeny (Boca Raton, FL: CRC Press, 2004), 348.
- Jacques Monod, quoted in Horace Freeland Judson, The Eighth Day of Creation: The Makers of the Revolution in Biology (New York: Simon & Schuster, 1979), 217.
Editor’s note: Dr. Wells’s latest book is Zombie Science: More Icons of Evolution. This article first appeared in Salvo 38. It is published here with the permission of Jonathan Wells.
Subscribe to:
Posts (Atom)