John11:34,35KJV"And said, Where have ye laid him? They said unto him, Lord, come and see.
35Jesus wept."
Why these tears for a saint who finally received his reward? If Jesus and his followers honestly believed that Lazarus was in heaven joyfully cavorting with the angels and saints in the presence of JEHOVAH God himself, would they not have responded quitedifferently to news of his departure from this life.
John11:24KJV"Martha saith unto him, I know that he shall rise again in the resurrection at the last day. " Note Martha's actual hope for her brother though.
where would she have gotten such an idea?From her Lord perhaps?
John6:39KJV"And this is the Father's will which hath sent me, that of all which he
hath given me I should lose nothing, but should raise it up again at the LAST DAY. "
No one goes to heaven when they die including Jesus himself. John20:17KJV"Jesus saith unto her, Touch me not; for I am not yet ascended to my
Father: but go to my brethren, and say unto them, I ascend unto my
Father, and your Father; and to my God, and your God."
Acts2:31KJV"He seeing this before spake of the resurrection of Christ, that his soul
was not left in hell, neither his flesh did see corruption." Thus like everyone elseJesus went to hell(sheol) when he died.His hope was his God and Father just like the rest of us.Hebrews5:7KJV"Who in the days of his flesh, when he had offered up prayers and
supplications with strong crying and tears unto him that was able to
save him from death, and was heard in that he feared;"
John11:34KJV"And said, Where have ye laid him? ..." Note please our Lord did not ask where have you laid his body but where have you laid HIM. Third person singularreferring to the person.obviously Lazarus was not in heaven.How could it be regarded as a kindness to recall anyone from the joy of heaven to the trials of this present age. Reject the mental contortions necessary to believe Christendom's falsehoods.
University of Bonn researchers think that they may have taught fish to count. They
tested the fact that many life forms can note the difference in small
quantities between “one more” and “one less,” at least up to five items,
on fish. Not much work had been done on fish in this area so they
decided to test eight freshwater stingrays and eight cichlids:
All of the fish were taught to
recognize blue as corresponding to “more” and yellow to “less.” The fish
or stingrays entered an experimental arena where they saw a test
stimulus: a card showing a set of geometric shapes (square, circle,
triangle) in either yellow or blue. In a separate compartment of the
tank, the fish were then presented with a choice stimulus: two gates
showing different numbers of shapes in the same color. When the fish
were presented with blue shapes, they were supposed to swim toward the
gate with one more shape than the test stimulus image. When presented
yellow shapes, the animals were supposed to choose the gate with one
less. Correct choices were rewarded with a food pellet. Three of the
eight stingrays and six cichlids successfully learned to complete this
task.
Rafael Núñez, a cognitive
scientist at the University of California, San Diego, who was not
involved in the study, regards the study as “well conducted,” adding
that “the problem is the interpretation.” For him, the paper provides
information about what he termed “quantical cognition” — the ability to
differentiate between quantities — in a 2017 paper. According to Núñez,
arithmetic or counting doesn’t have to be invoked to explain the results
in the present paper. “I could explain this result by . . . a fish or
stingray having the perceptual ability to discriminate quantities: in
this case, this will be to learn how to pick, in the case of blue, the
most similar but more, and in the case of yellow, the most similar but
less. There’s no arithmetic here, just more and less and similar.”
The problem, as Núñez says, is with interpretation. Animal cognition researcher Silke Goebelpoints out that
many life forms can distinguish between “more” and “less” in large
numbers. Researchers have also found that, so far, infants, fish, and
bees can recognize changes in number between 1 and 3. But they don’t get
much beyond that.
To say seriously that fish “do math” would, of course, be misleading.
Mathematics is an abstract enterprise. The same operations that work
for single digits work for arbitrarily large numbers. It is possible to
calculate using infinite (hyperreal) numbers. There are imaginary numbers,unexplained/unexplainable numbers, and at least one unknowable number. But we are stepping out into territory here that will not get a fish its food pellet.
Still, it’s a remarkable discovery that many life forms can
manipulate quantities in a practical way. Here are some other recent
highlights.
Read the rest at Mind Matters News, published by Discovery Institute’s Bradley Center for Natural and Artificial Intelligence.
We’ve just ended the first quarter of the year. It’s a long way to New Year’s Eve 2022. But this new open access paper from
senior author Sara Walker (Arizona State) and her collaborators will be
hard to top, in the “Wow, that is so interesting!” category. (The first
author of this paper is Dylan Gagler, so we’ll refer to it as “Gagler et al. 2022” below.)
1. Back in the day, the best evidence for a single Tree of
Life, rooted in the Last Universal Common Ancestor (LUCA), was the
apparent biochemical and molecular universality of Earth life.
Leading neo-Darwinian Theodosius Dobzhansky expressed this point eloquently in his famous 1973 essay, “Nothing in biology makes sense except in the light of evolution”:
The unity of life is no less
remarkable than its diversity…Not only is the DNA-RNA genetic code
universal, but so is the method of translation of the sequences of the
“letters” in DNA-RNA into sequences of amino acids in proteins. The same
20 amino acids compose countless different proteins in all, or at least
in most, organisms. Different amino acids are coded by one to six
nucleotide triplets in DNA and RNA. And the biochemical universals
extend beyond the genetic code and its translation into proteins:
striking uniformities prevail in the cellular metabolism of the most
diverse living beings. Adenosine triphosphate, biotin, riboflavin,
hemes, pyridoxin, vitamins K and B12, and folic acid implement metabolic
processes everywhere. What do these biochemical or biologic
universals mean? They suggest that life arose from inanimate matter only
once and that all organisms, no matter now diverse, in other respects,
conserve the basic features of the primordial life.[Emphasis added.]
For Dobzhansky, as for all neo-Darwinians (by definition), the
apparent molecular universality of life on Earth confirmed Darwin’s
prediction that all organisms “have descended from some one primordial
form, into which life was first breathed” (1859, 494) — an entity now
known as the Last Universal Common Ancestor, or LUCA. So strong is the
pull of this apparent universality, rooted in LUCA, that any other
historical geometry seems unimaginable.
The “Laws of Life”
Theoretician Sara Walker and her team of collaborators, however, are looking for an account of what they call (in Gagler et al. 2022)
the “laws of life” that would apply “to all possible biochemistries” —
including organisms found elsewhere in the universe, if any exist. To
that end, they wanted to know if the molecular universality explained
under neo-Darwinian theory as material descent from LUCA (a) really
exists, and (b) if not, what patterns do exist, and how might those be explained without presupposing a single common ancestor.
And a single common ancestor, LUCA? That’s what they didn’t find.
2. Count up the different enzyme functions — and then map that number within the total functional space.
Many thousands of different enzyme functional classes,
necessary for the living state, have been described and catalogued in
the Enzyme Commission Classification, according to their designated EC
numbers. These designators have four digits, corresponding to
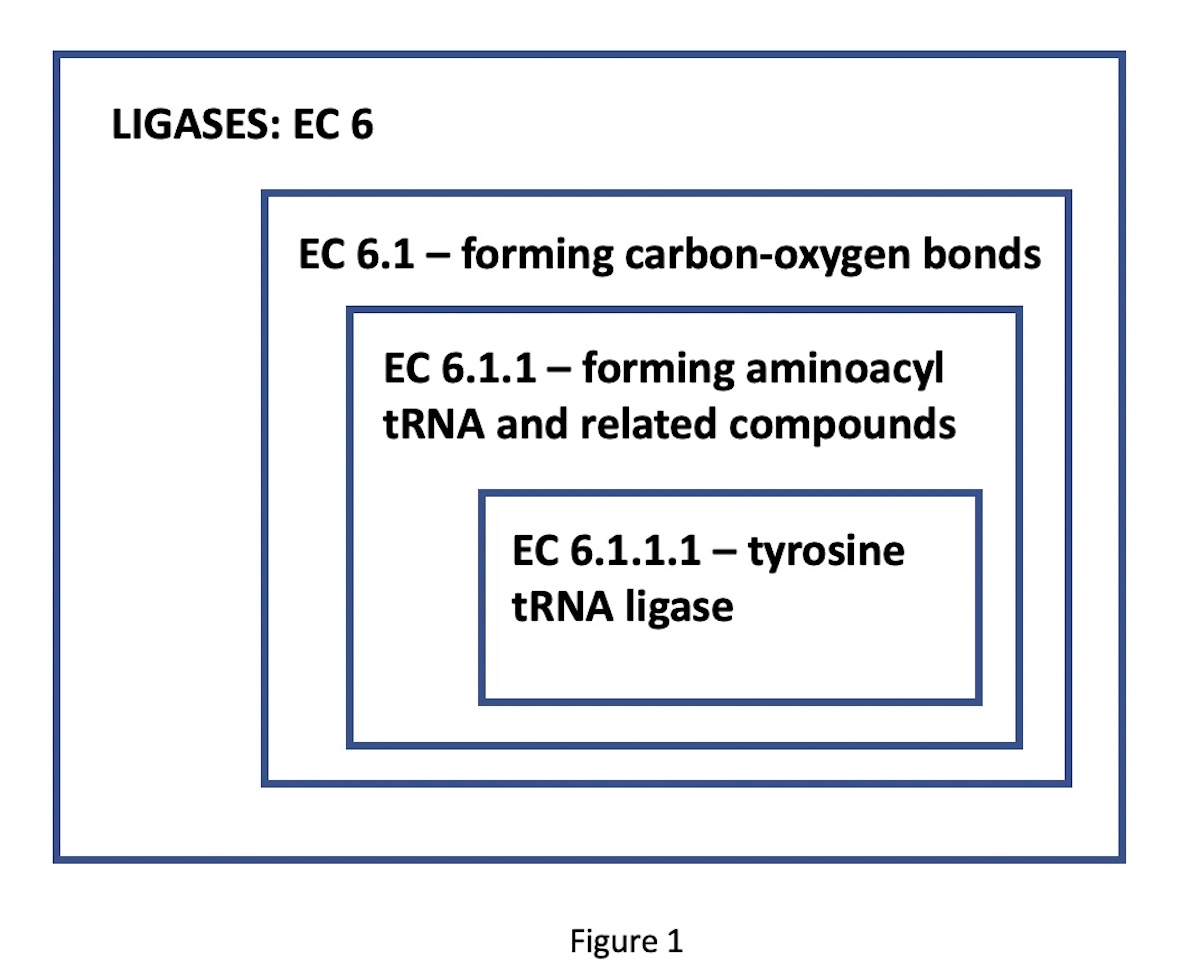
progressively more specific functional classes. For instance, consider
the enzyme tyrosine-tRNA ligase. Its EC number,
6.1.1.1, indicates a nested set of classes: EC 6 comprises the ligases
(bond-forming enzymes); EC 6.1, those ligases forming carbon-oxygen
bonds; 6.1.1, ligases forming aminoacyl-tRNA and related compounds;
finally, 6.1.1.1, the specific ligases forming tyrosine tRNA. (See
Figure 1.)
The Main Takeaway from This Pattern?
Being a ligase — namely, an enzyme that forms bonds using ATP — entails belonging to a functional group, but not a group with material identity among its members.
A rough parallel to a natural language such as English may be helpful.
Suppose you wanted to express the idea of “darkness” or “darkened”
(i.e., the relative absence of light). English supplies a wide range of
synonyms for “darkened,” such as:
murky
shaded
shadowed
dimmed
obscured
The same would be the case — the existence of a set of synonyms,
i.e., words with the same general meaning, but not the same sequence
identity — for any other idea. The concept of something being “blocked,”
for instance, takes the synonyms:
jammed
occluded
prevented
obstructed
hindered
While these words convey (approximately) the same meaning, and hence fall into the same semantic functional classes, they are not the same character strings.
Their locations in an English dictionary, ordered by alphabet sequence,
may be hundreds of pages apart. Moreover, as studied by the discipline
of comparative philology, the historical roots of a word such as
“hindered” will diverge radically from its functional synonyms, such as
“blocked.” These two words, although semantically largely synonymous,
enter English from originally divergent or unrelated antecedents — a
character string gap still reflected by their very different spellings.
A strikingly similar pattern obtains with the critical (essential) components of all organisms. Gagler et al. 2022
looked at the abundances of enzyme functions across the three major
domains of life (Bacteria, Archaea, Eukarya), as well as in metagenomes
(environmentally sampled DNA). What they found was remarkable — a
finding (see below) which may be easier for non-biological readers to
understand via another analogy.
3. A segue into computer architectures — then back to enzymes.
The basic architecture of laptop computers includes components present in any such machine, defined by their functional roles:
Central processing unit (CPU) — the primary logic operator
Memory — storage of coded information
Power supply — electrons (energy) needed for anything at all to be computed
And so on. (Although exploring this point in detail would take us far
afield, it is worth noting that in 1936, when Alan Turing defined a
universal computational machine, he did so with no idea about the
arrival, decades down the road, of silicon-based integrated circuits,
miniaturized transistors, motherboards, solid-state memory devices, or
any of the rest of the material parts of computers now so familiar to us. Rather, his parts were functionally, not materiallydefined, asabstractions
occupying the various roles those parts would play in the computational
process — whatever their material instantiation would later turn out to
be.) Now suppose we examined 100,000 laptops, randomly sampled from
around the United States, to see what type of CPU — meaning which materialpart (e.g., built by which manufacturer) — each machine used as its primary logic operator.
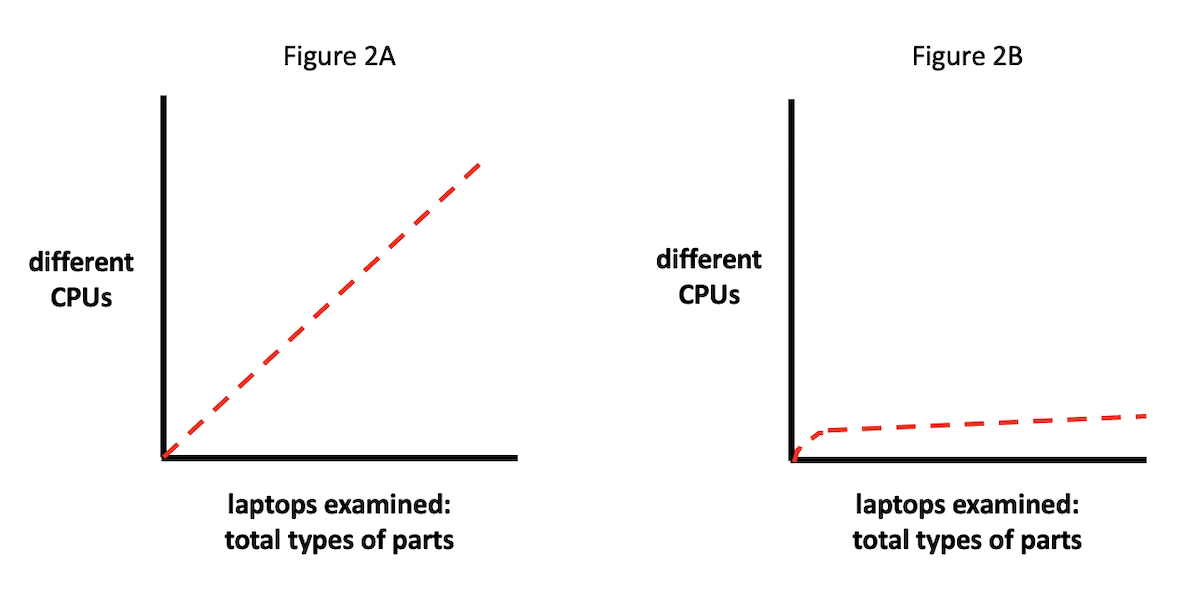
A range of outcomes is possible (see Figures 2A and 2B). For
instance, if we plot CPUs from different manufacturers on the y axis,
against the total number of laptop parts inspected on the x axis, it
might be the case that the distribution of differently manufactured
(i.e., materially distinct) CPUs would scale linearly with laptops
inspected (Figure 2A). In other words, as our sample of inspected laptop
parts grows, the number of different CPUs discovered would trend
upwards correspondingly.
Or — and this fits, of course, with the actual situation we find (see
Figure 2B) — most of the laptops would contain CPUs manufactured either
by Intel or AMD. In this case, we would plot a line whose slope would
change much more slowly, staying largely flat, in fact, after the CPUs
from Intel and AMD were tallied.
The Core Rationale of Their Approach
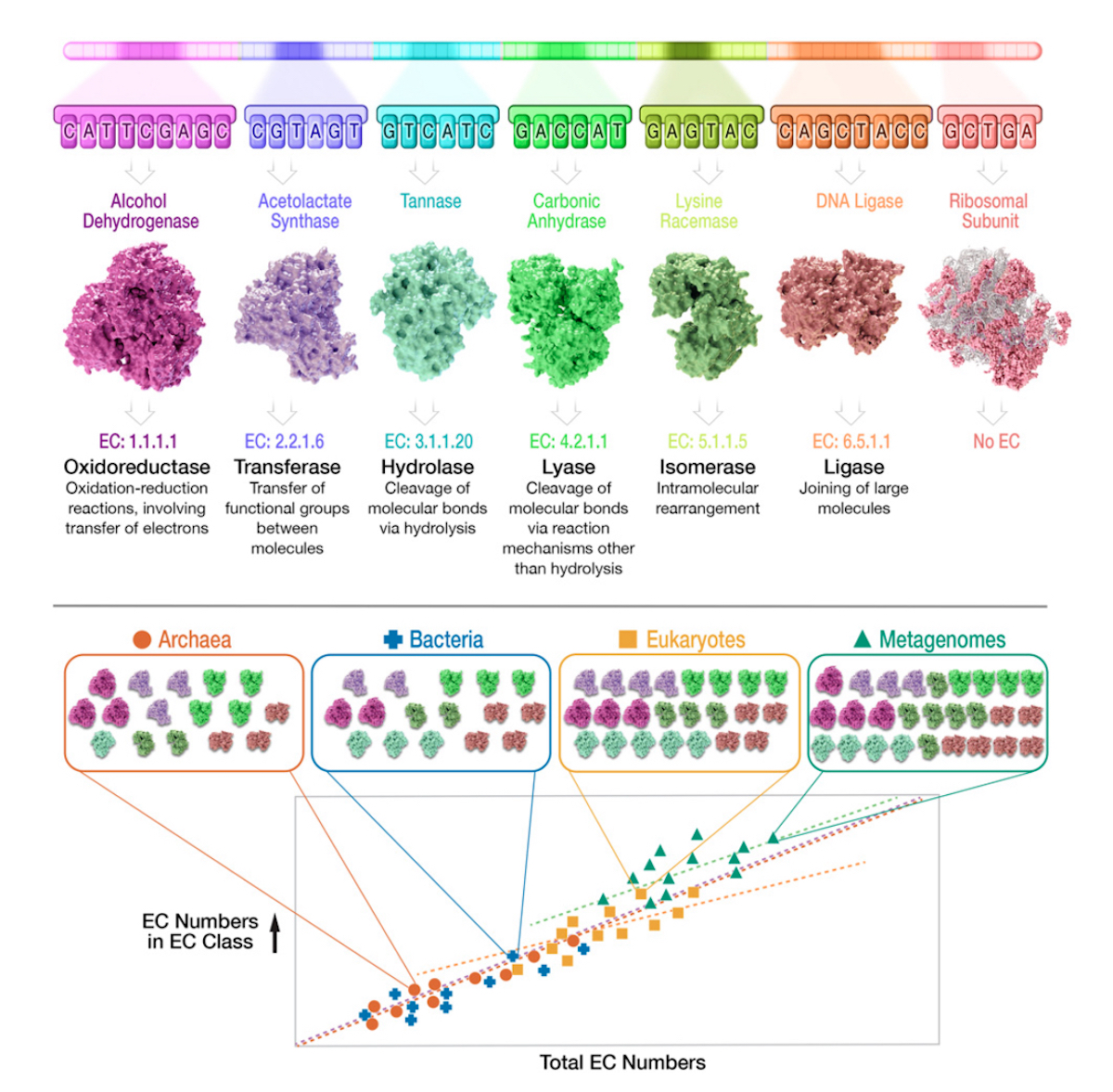
Now consider Figure 3 (below), from the Gagler et al. 2022
paper. This shows the core rationale of their approach: tally the
EC-classified enzyme “parts” within each of the major domains, and from
metagenomes, and then plot that tally against the total EC numbers.
Figure 3 is used from Gagler et al. 2022 under Creative Commons License 4.0 (CC BY-NC-ND).
Figure 3 also shows their main finding. As the enzyme reaction space
grows (on the horizontal axis — total EC numbers), so do the number of
unique functions (on the vertical axis — EC numbers in each EC class).
The lesson that Gagler et al. 2022 draw from this discovery? The pattern is NOT due to material descent from a single common ancestor, LUCA. Indeed,
under the heading, “Universality in Scaling of Enzyme Function Is Not
Explained by Universally Shared Components,” they explain that material
descent from LUCA would entail shared “microscale features,” meaning
“specific molecules and reactions used by all life,” or “shared
component chemistry across systems.” If we use the CPU / laptop analogy,
this microscale commonality would be equivalent to finding CPUs from
the same manufacturer, with the same internal logic circuits, in every
laptop we examine.
But what Gagler et al. 2022 found was a macroscale pattern,
“which does not directly correlate with a high degree of microscale
universality,” and “cannot be explained directly by the universality of
the underlying component functions.” In an accompanying news story,
project co-author Chris Kempes, of the Santa Fe Institute, described
their main finding in terms of functional synonyms: macroscale functions
are required, but not the identical lower-level components:
“Here we find that you get these
scaling relationships without needing to conserve exact membership. You
need a certain number of transferases, but not particular transferases,”
says SFI Professor Chris Kempes, a co-author on the paper. “There are a
lot [of] ‘synonyms,’ and those synonyms scale in systematic ways.”
As Gagler et al. frame the point in the paper itself (emphasis added):
A critical question is whether the
universality classes identified herein are a product of the shared
ancestry of life. A limitation of the traditional view of biochemical
universality is that universality can only be explained in terms of
evolutionary contingency and shared history, which challenges our
ability to generalize beyond the singular ancestry of life as we know
it. …Instead, we showed here that universality classes are not directly correlated with component universality,
which is indicative that it emerges as a macroscopic regularity in the
large-scale statistics of catalytic functional diversity. Furthermore, EC universality cannot simply be explained due to phylogenetic relatedness since the range of total enzyme functions spans two orders of magnitude, evidencing a wide coverage of genomic diversity.
Sounds Like Intelligent Design
It is interesting to note that this paper was edited (for the PNAS)
by Eugene Koonin of the National Center for Biotechnology Information.
For many years, Koonin has argued in his own work that the putative
“universality due to ancestry” premise of neo-Darwinian theory no longer
holds, due in large measure to what he and others have termed
“non-orthologous gene displacement” (NOGD). NOGD is a pervasive pattern
of the use of functional synonyms — enzyme functions being carried out
by different molecular actors — in different species. In 2016, Koonin wrote:
As the genome database grows, it
is becoming clear that NOGD reaches across most of the functional
systems and pathways such that there are very few functions that are
truly “monomorphic”, i.e. represented by genes from the same orthologous
lineage in all organisms that are endowed with these functions.
Accordingly, the universal core of life has shrunk almost to the point
of vanishing…there is no universal genetic core of life, owing to the
(near) ubiquity of NOGD.
Universal functional requirements, but without the identity of material components — sounds like design.
Wow, the new Long Story Short video is out now, and I think
it’s the best one yet — it’s amazingly clear and quite funny. You’ll
want to share it with friends. Some past entries in the series have
considered the problems associated with chemical evolution, or
abiogenesis, how life could have emerged from non-life on the early
Earth without guidance or design. The new video examines cell membranes,
which some might imagine as little more than a soap bubble or an
elastic balloon. This is VERY far from the case.
To keep the cell alive, there’s an astonishing number of complex and
contradictory things a cell membrane needs to do. If unassisted by
intelligent design, how did the very first cell manage these tricks?
It’s a puzzle, since “The membrane had to be extremely complex from the
very BEGINNING, or life could never begin.” Some materialists have an
answer: protocells, a simpler version of the simplest cells we know of
today. But, asks Long Story, could a necessarily fragile,
simpler cell survive without assistance from its environment, something
like a hospital ICU? It seems not. If so, that makes any unguided
scenario of abiogenesis a non-starter. We’ll have more to say in coming
days about the science behind this.
The latest video in the Long Story Short series was released
this week on YouTube. The video explains how cell membranes in all of
life display complexity that cannot be explained by purely natural
processes. See my comments from yesterday, “New Animated Video: Cell Membranes by Natural Processes Alone?,” adding some supporting details to the argument. Here’s more.
As we uncover layer after layer of the astounding complexity of even
the simplest forms of life, the origin-of-life research community
increasingly relies upon their trump card: imaginary protocells that
supposedly existed long ago and were dramatically simpler than existing
life. As the story goes, modern life may indeed be very complex, but
protocells used to be much simpler, and there was plenty of time for the
complexity to develop.
Protocells conveniently fill the uncomfortably large gap between the
simple molecules that can be produced by prebiotic processes and the
staggering complexity of all extant life. But there are three major
problems with the concept of protocells. These problems are all backed
by strong empirical support, in sharp contrast with the concept of
protocells.
A Coddling Environment
First, scientists have been working for decades to simplify existing
life, trying to arrive at a minimal viable life form by jettisoning
anything that is not essential from the simplest extant cells. The
success of Craig Venter’s group is well known. Building on their efforts
to produce synthetic life (“Synthia” or “Mycoplasma labritorium”) in 2010,1,2 in 2016 they introduced the current record holder for the simplest autonomously reproducing cell (JVCI Syn3.0).3 With
a genome of only 473 genes and 520,000 base pairs of DNA, JVCI Syn3.0
can reproduce autonomously, but it certainly isn’t robust. Keeping it
alive requires a coddling environment — essentially a life-support
system. To arrive at a slightly more stable and robust organism that
reproduced faster, the team later added back 19 genes to arrive at JVCI
Syn3A.4 When combined, this work provides an approximate
boundary for the simplest possible self-replicating life. We are clearly
approaching the limit of viable cell simplicity. It seems safe to
conclude that at least 400 genes (and approximately 500,000 base pairs
of DNA) are the minimum requirements to produce a self-replicating
cell.
Exporting to the Environment
Second, we know that the process of simplifying an existing cell by
removing some of its functionality doesn’t actually simplify the overall
problem — it only exports the required complexity to the environment. A
complex, robust cell can survive in changing conditions with varying
food sources. A simplified cell becomes dependent on the environment to
provide a constant, precise stream of the required nutrients. In other
words, the simplified cell has reduced ability to maintain homeostasis,
so the cell can only remain alive if the environment takes on the
responsibility for homeostasis. Referring to JVCI Syn3A, Thornberg et al. conclude,
“Unlike most organisms, which have synthesis pathways for most of
[their] building blocks, Syn3A has been reduced to the point where it
relies on having to transport them in.”5 This implies that
the environment must provide a continuous supply of more specific and
complex nutrients. The only energy source that JVCI Syn3A can process is
glucose,4 so the environment must provide a continuous
supply of its only tolerable food. Intelligent humans can provide such a
coddling life-support environment, but a prebiotic Earth could not.
Protocells would therefore place untenable requirements on their
environment, and the requirements would have to be consistently met for
millions of years.
Striving for Simplicity
Third, we know that existing microbes are constantly trying to
simplify themselves, to the extent that their environment will allow. In
Richard Lenski’s famous E. coli experiment, the bacteria
simplified themselves by jettisoning their ribose operons after a few
thousand generations, because they didn’t need to metabolize ribose and
they could replicate 2 percent faster without it, providing a selective
advantage.6 Furthermore, Kuo and Ochman studied the
well-established preference of prokaryotes to minimize their own DNA,
concluding: “deletions outweigh insertions by at least a factor of 10 in
most prokaryotes.”7 This means that existing life has been
trying from the very start to be as simple as possible. Therefore, it is
likely that extant life has already reached something close to the
simplest possible form, unless experimenters like Lenski provide a
coddling environment for a long duration that allows further
simplification. But such an environment requires the intervention of
intelligent humans to provide just the right ingredients, at the right
concentrations, and at the right time. No prebiotic environment could do
this. Therefore, scientists need not try to simplify existing life — we
already have good approximations of the simplest form. Indeed, Mycoplasma genitalium has a genome of 580,000 base pairs and 468 genes8 whereas Craig Venter’s minimal “synthetic cell” JVCI Syn3.0 has a comparable genome of 520,000 base pairs and 473 genes.3
The data provide a clear picture: the surprising complexity of even
the simplest forms of existing life — 500,00 base pairs of DNA — cannot
be avoided and cannot be reduced unless intelligent agents provide a
complex life-support environment. Because protocells would have had to
survive and reproduce on a harsh and otherwise lifeless planet,
protocells are not a viable concept. Protocells place origin-of-life
researchers in a rather awkward position: relying upon an imaginary
entity to sustain their belief that only matter and energy exist.
References
Gibson DG et al. Creation of a bacterial cell controlled by a chemically synthesized genome. Science 2010; 329:52–56.
Thornburg ZR et al. Fundamental behaviors emerge from simulations of a living minimal cell. Cell 2022; 185: 345-360.
Cooper VS et al. Mechanisms causing rapid and parallel loss of ribose catabolism in evolving populations of Escherichia coli B. J Bacteriology 2001, 2834-2841.
Kuo, CH and Ochman H. Deletional bias across the three domains of life. Genome. Biol. Evol. 1:145–152.
Fraser CM et al. The minimal gene complement of Mycoplasma genitalium. Science. 1995; 270; 397-403.