Saturday, 30 April 2022
China vs. China?
<iframe width="853" height="480" src="https://www.youtube.com/embed/6FHHT_O-Khk" title="YouTube video player" frameborder="0" allow="accelerometer; autoplay; clipboard-write; encrypted-media; gyroscope; picture-in-picture" allowfullscreen></iframe>
MAGA: Africa edition?
<iframe width="853" height="480" src="https://www.youtube.com/embed/GHDwLP9WLQI" title="YouTube video player" frameborder="0" allow="accelerometer; autoplay; clipboard-write; encrypted-media; gyroscope; picture-in-picture" allowfullscreen></iframe>
Urban nomad.
<iframe width="853" height="480" src="https://www.youtube.com/embed/B_UZyT0nShI" title="YouTube video player" frameborder="0" allow="accelerometer; autoplay; clipboard-write; encrypted-media; gyroscope; picture-in-picture" allowfullscreen></iframe>
Dembski on being an I.D. pioneer.
ID Pioneer William Dembski on His Rocky and Rewarding Journey
On a new episode of ID the Future, intelligent design pioneer William Dembski tells the story of his rocky journey into and out of higher education, the reasons for his sabbatical from the ID movement, his recent success as an entrepreneur, and his return to ID work. Along the way Dembski bats down a mistaken rumor about his sabbatical. The occasion for his conversation with host Casey Luskin is the recent anthology Dembski and Luskin contributed to and helped edit, The Comprehensive Guide to Science and Faith: Exploring the Ultimate Questions about Life and the Cosmos. Download the podcast or listen to it here.
And still yet more primeval tech vs. Darwin.
<iframe width="770" height="433" src="https://www.youtube.com/embed/9qOOeAzKOoY" title="YouTube video player" frameborder="0" allow="accelerometer; autoplay; clipboard-write; encrypted-media; gyroscope; picture-in-picture" allowfullscreen></iframe>
David Berlinski is skeptical about the skeptics?
Medved, Berlinski Take on Steven Pinker and Whig History
On a new episode of ID the Future, Human Nature author and polymath David Berlinski and radio host Michael Medved discuss everything from human depravity, the burning of Notre Dame, and the Russian invasion of Ukraine to the Big Bang and a quixotic century-old pact to ban war. Download the podcast or listen to it here.
Berlinski argues that the case for the death of God and the case for
the impending demise of human depravity have been greatly exaggerated.
Taking issue with Steven Pinker, Berlinski insists that there is little
if any evidence that human evil is being steadily rolled back by the
spread of secular values. Further, the idea that science has disproven
God flies in the face of trends running in the opposite direction,
perhaps most dramatically in the triumph of the Big Bang theory over an
eternal universe model. Berlinski, who himself is not religious, insists
that optimistic Whig history is bankrupt and that anyone imagining that
human depravity and the God hypothesis are things of the past are
themselves living in the past.
Wednesday, 27 April 2022
On the myth of engineerless engineering.
Unraveling the Myth that Undesigned Processes Generate Novel Functions
I was recently informed of a video posted on the YouTube channel Creation Myths titled “Creation Myth: ‘Information’ Requires ‘Intelligence’.” The video specifically references a clip of Stephen Meyer detailing the design implications of the Cambrian Explosion. Meyer states that the information required for the sudden appearance of radically new animals could only have originated from a mind. The critic responds to this claim by arguing that experiments have demonstrated that information can be created by natural processes, and he cites two research studies to support this assertion. His argument ultimately fails since it is founded on a misunderstanding of the evidence for design associated with biological information. This error is so common that it deserves special attention.
The Research Studies
The first cited article is a 2017 study by Neme et al. that purportedly demonstrated the creation of new information with ease. The researchers inserted randomly generated sequences of 150 base pairs into the DNA of E. coli. They reported that 25 percent of random sequences enhance cells’ growth rate. The experiment purportedly yielded new information without intelligent direction.
The second cited article is a 2018 study by Yona et al. that explored the difficulty of randomly generating a 100-base-pair DNA sequence in E. coli that would bind to an RNA polymerase. The study demonstrated that 10 percent of random sequences adjacent to the genes in a lac operon would
bind to the polymerase in such a way as to initiate transcription. This
study also purportedly demonstrates that information can be created by a
random process.
The Misunderstanding
Upon close inspection, both studies fail to challenge the design argument that is based on biological information. Neme et al. misinterpreted their results, as Weisman and Eddy explain in their critical review of the study. Douglas Axe summarizes the experimenters’ error as follows:
They merely showed that if you burden bacteria by forcing them to churn out RNA and protein from random inserts, it’s fairly easy to find sequence-dependent effects on growth — not because anything clever has been invented, but because the burden of making so much junk varies slightly with the kind of junk. That means any junk that slows the process of making more junk by gumming up the works a bit would provide a selective benefit. Such sequences are “good” only in this highly artificial context, much as shoving a stick into an electric fan is “good” if you need to stop the blades in a hurry.
In short, the sequences performed no new function, so no new information was created.
The Yona et al. experiment did show that a DNA sequence can be randomly generated that can perform simple functions, such as binding to a polymerase. Yet this achievement is not relevant to Stephen Meyer’s full argument. Meyer is not claiming that random processes cannot generate small quantities of information. He is arguing that random processes cannot generate the quantity of information required for anything comparable to creating a new protein with a novel structure. Axe and others have decisively demonstrated that the information associated with even modest proteins is typically greater than what could be produced by any undirected process (here, here, here).
The Challenge for Evolution
The central challenge for evolutionary theory is creating sufficient
information to produce something truly novel that functions at a level
that would benefit an organism. In the case of the lac operon,
the specificity required for it to function is not the difficulty of an
RNA polymerase binding to the promoter region. The specificity and thus
the information reside in the sequences that encode the repressor that
acts as an on/off switch and the genes that encode the proteins that
break down lactose. The minimal required information for the operon to
function is vastly greater than that associated with the region that
binds to a polymerase. The amount is almost certainly beyond what any
undirected process could produce.
On the History of life's Barrage against Darwinism
Bechly: “Life’s Second ‘Big Bang'”
German paleontologist Günter Bechly is co-author (with Stephen Meyer) of the chapter titled “The Fossil Record and Universal Common Ancestry” in the book Theistic Evolution: A Scientific, Philosophical, and Theological Critique. On a classic episode of ID the Future with host Sarah Chaffee, he moves on from the Cambrian explosion (see here) to discuss “life’s second ‘big bang.’” He then touches on other biological explosions, including the Avalon explosion, the Triassic explosion, the origin of flowering plants, and the origin of placental mammals. “There’s no reasonable way,” Bechly concludes, “to get from bacteria to mammals via evolutionary processes.” Download the podcast or listen to it here.
Darwinism in time trouble?
Species Pairs: A New Challenge to Darwinists
In the many years of vehement debate between proponents of unguided evolution and intelligent design, it sometimes may look like all has already been said and there are no new arguments on either side. However, this is not the case at all. Intelligent design theory has greatly developed since its early beginnings and many new arguments have been added in support of the design inference. Here, I want to introduce another new argument and formulate a challenge to my Darwinist colleagues. This challenge is by no means rhetorical and could be easily met with simple research in publicly available data bases. Here it is.
As I have laid out in various publications (e.g., Bechly & Meyer 2017) and lectures, the fossil record demonstrates that the history of life was not a series of gradual transformations by an accumulation of small changes over long periods of time. Instead of conforming to this gradualist prediction of Darwin’s theory of evolution, the fossil record consistently documents a series of saltational transitions with abrupt appearances of new body plans within very short windows of time. This implies a fatal problem for Darwinism called the waiting time problem, because population genetic calculations and simulations show that the windows of time established by the fossil record are orders of magnitude too short to accommodate the required genetic changes for these body plan transformations.
Some examples of abrupt body plan transitions are the origin of photosynthesis; the origin of eukaryotes; the origin of the Ediacaran biota (Avalon Explosion) and Cambrian animal phyla (Cambrian Explosion) such as the origin of trilobites from worm-like ancestors in less than 13 million years (Daley et al. 2018, Bechly 2018); the origin of efficient eyes in arthropods, cephalopods, and vertebrates; the terrestrialization of plants (embryophytes), arthropods (tracheae), and vertebrates (tetrapod limbs); the origin of wings in insects, pterosaurs, bats, and birds (including the origin of pennaceous feathers from filamentous precursors); the origin of secondarily marine vertebrates such as ichthyosaurs, mosasaurs, manatees, and whales; the origin of echolocation in bats and whales; the origin of complex new reproductive systems (angiosperm flowers, dragonfly secondary copulatory apparatus, holometabolic insect metamorphosis, amniote egg, and eutherian placenta); the origin of distinct new body plans in vertebrates (e.g., snakes, turtles, bats, and whales); and even the origin of our own genus Homo and of a globular braincase correlated with the “Creative Explosion” of symbolic thinking within Homo sapiens.
The Lifespan of a Single Species
An additional fact that underscores the waiting time problem in cases like whale origins (see this Long Story Short 2020 video) is the average longevity of larger mammal species and especially artiodactyls at only 4.1-4.39 million years (Prothero 2014). Whales are thought to be nested in and derived from artiodactyl ungulates. The fossil record shows that the transition from quadrupedal whale ancestors similar to Raoellidae (such as Indohyus) and Pakicetidae to fully marine pelagicete whales like Basilosauridae happened in just 4.5 million years. This implies that the body plan transition from a pig-like animal to a dolphin-like animal happened within the lifespan of a single species. Of course, this does not exclude the possibility of several successive speciation events within this time period, but it is still a fact that illustrates the biological abruptness of this major anatomical re-engineering.
Finally, there is also another related problem that has been hitherto largely overlooked: The morphological similarity of modern species pairs, which have diverged in a similar time frame, poses a severe problem. That is because it implies that the macroevolutionary processes that allegedly were at work and common during all periods of Earth history and in all groups of organisms, apparently were totally absent in the origins of all of the millions of living species. To explore this issue, I surveyed TimeTree.org (Hedges & Kumar 2009, Hedges et al. 2006, 2015, Kumar et al. 2017), which is a databank of 97,085 living species of different groups of organisms, with molecular clock estimates of their time of divergence based on 3,998 studies. When probing any pairs of species, even those with longer divergence times than available for the development of the body plan differences between pakicetids and basilosaurids, we find without exception that their morphologies are hardly distinguishable for laymen and they often still can hybridize.
So Much for the Theoretical Prelude
Now let’s look at some more-or-less random examples from TimeTree to see what I mean.
Firs (Abies spec.) and cedars (Cedrus spec.) belong to the same subfamily of conifer trees but separated already 141 million years ago. That is more than 30 times the time available for the origin of marine whales, but there is hardly any difference in body plan. Of course, this might just be a case of evolutionary stasis as in living fossils, but the next examples will make you think again.

{kind=link}

{kind=link}
The common house fly (Musca domestica) and the small house fly (Fannia scalaris) diverged about 48 million years ago. For laymen they look basically indistinguishable. Hmmm, that’s strange, but could still be a fluke.

{kind=link}

{kind=link}
My favorite animals are dragonflies and damselflies. The northern damselfly (Coenagrion hastulatum) and the azure damselfly (Coenagrion puella) diverged 11.8 million years ago. Even as an expert on these insects, I have to resort to a determination key to distinguish them. Can you?


.jpg){kind=link}
What about amphibians? The European common frog (Rana temporaria) and moor frog (Rana arvalis) diverged 21.4 million years ago. They look almost identical. Do we start to see a pattern?

{kind=link}

{kind=link}
“Reptiles” show the same pattern. The Galapagos land iguanas (Conolophus spec.) and marine iguanas (Amblyrhynchus spec.) diverged 18.2 million years ago. The marine iguanas can excrete salt from a gland at their nostrils and have a more flattened tail, but otherwise still look very much like their cousins.

{kind=link}

{kind=link}
Avian Examples
Birds rank among the best studied groups in terms of speciation. The green warbler (Phylloscopus nitidus) and Bonelli’s warbler (P. bonelli) diverged 15.2 million years ago (but at least 4-7 million years according to Helbig et al. 1995). They look identical and may still be able to hybridize.

_(Green_Warbler),_Chorokhi_Delta,_Georgia.jpg){kind=link}

{kind=link}
Here is another avian example to show this is a common pattern among recent bird species. The house sparrow (Passer domesticus) and tree sparrow (Passer montanus) diverged 10.2 million years ago. They have some minor differences in color pattern but indeed hybridize even in the wild. Still thinking evolution can achieve miracles in a few million years? Where the heck is the evidence?

.jpg){kind=link}

{kind=link}
Well, let’s move on to mammals. The common house mouse (Mus musculus) and house rat (Rattus rattus) diverged 20.9 million years ago (at least 12 million years according to Kimura et al. 2015). Apart from the size difference they look very much alike.

{kind=link}

{kind=link}
Did you ever taste beefalo steak? Beefalos are hybrids between European bison (Bison bonasus) and cattle (Bos taurus) that diverged 4.88 million years ago, which is about the same time frame as in the whale example. Some archaic breeds of cattle like the Scots highland cattle look even more similar to the bison and the aurochs.

.jpg){kind=link}

{kind=link}
Horse (Equus caballus) and ass (Equus asinus) diverged 7.7 million years ago and can still hybridize as mules. Their wild ancestors looked even more similar than most of their modern domesticated breeds.

{kind=link}

{kind=link}
Asian elephants (Elephas maximus) and African elephants (Loxodonta africana) diverged 25.9 million years ago (at least 7.6 million years according to Rohland et al. 2007), and they mainly differ in the tip of the trunk, ear size, and shape of their withers. Even the two almost identical African species of the savannah elephant (Loxodonta africana) and the forest elephant (Loxodonta cyclotis) diverged 7.6 million years ago (at least 4 million years according to Rohland et al. 2007). Not really a new body plan.

.jpg){kind=link}

{kind=link}

{kind=link}
The South American spectacled bear (Tremarctos ornatus) and Asian black bear (Ursus / Selenarctos thibetanus) diverged 16.5 million years ago. They look very similar and can hybridize in captivity (Mondolfi & Boede 1981).

{kind=link}

{kind=link}
River otter (Lutra lutra) and brown fur seal (Arctocephalus pusillus) diverged about 40 million years ago. They indeed look quite different, but still a far cry from the difference between pakicetids and basilosaurids in a tenth of the time. This example is interesting because some experts thought that otters represent the closest related group to pinnipeds, or at least assumed stem pinnipeds like the Miocene Puijila darwini were very similar to otters. Therefore, this case seems to be a pretty good analogue to the early amphibious stem whales, which made a much bigger transition in terms of body plan in just a few million years.

{kind=link}

{kind=link}
The Sister Group of Whales
In modern phylogenetic reconstructions hippos are consistently recovered as the sister group of whales. So, we might expect to find a comparable evolutionary disparity within this group. However, even though the river hippo (Hippopotamus amphibius) and pygmy hippo (Choeropsis liberiensis) diverged 9.6. million years ago, they share the same body plan and only differ somewhat in size and proportions. Still not convinced?

{kind=link}

{kind=link}
Now, let’s have a look at modern cetaceans. Maybe they are different? The common dolphin (Delphinus delphis) and the bottlenose dolphin (Tursiops truncatus) diverged 3.99 million years ago. This represents about the timeframe available between pakicetids and basilosaurids and shows what blind evolution at best can achieve with whales in this time: only very minor differences!

_(14825276609).jpg){kind=link}

{kind=link}
Finally, what about great apes and humans. Chimp (Pan paniscus) and gorilla (Gorilla gorilla) diverged according to TimeTree 9.06 million years ago and humans (Homo sapiens) from chimps 6.7 million years, which agrees with the hominin fossil record. There are two possibilities: Either you follow those scientists who consider the biological difference between humans and chimps as marginal. Then this example would just confirm the pattern described above. Or, you consider humans as very different from chimps, based on their different bipedal locomotion and especially their mental capacity and cultural achievements. In the latter case humans would represent the only exception to the pattern that I could find, which would be a remarkable confirmation of Judeo-Christian human exceptionalism.

.jpg){kind=link}

{kind=link}

{kind=link}
Two Indisputable Facts
These examples could be expanded endlessly but should be sufficient to establish the point. There are clearly limits to what unguided evolution can do within a few million years, and these limits are far below the level of any major body plan transitions. Thus, we can safely conclude that there are two indisputable facts that require an adequate explanation:
1.) There are many examples of fossil species pairs with very different body plans that diverged within a window of time of 5 (±5) million years. This is even more remarkable if we consider that there are only about 350,000 described fossil species (extrapolated based on data in Teichert 1956, Valentine 1970, Raup 1976, and Alroy 2002), which represent only a tiny fraction of the estimated 5-50 billion species that have ever lived on Earth (Raup 1991).
2.) There exist no living species pairs with even remotely similar differences in body plan that are dated to have diverged in a similar time frame. This is even more remarkable if we consider that there are an estimated 8.7 million living species (Mora et al. 2011, Strain 2011, Sweetlove 2011), of which more than 2 million are described (IISE 2012). Previous estimates of the total number of living species varied from 3-100 million species (May 1988, Tangley 1997, Chapman 2009), but if microbes are included, it could even be up to a trillion living species (Locey & Lennon 2016, Latty & Lee 2019).
Considering the fact that windows of time of only 5-10 million years account for most of the abrupt appearances of new body plans in the fossil record (Bechly & Meyer 2017, Bechly 2021), the Bayesian likelihood of not finding a single example of similar morphological disparity having originated on a similar time frame among the millions of living species is basically close to zero. I consider this simple argument as a final nail in the coffin of Darwinian unguided evolution.
A Public Challenge
Having made my case, I here formally and publicly pose the challenge again to prove me wrong. My dear Darwinist friends and colleagues, please find in the vast database of 97,000 species at TimeTree.org just a single example of any pair of different species that have diverged about 5 million years ago (give or take a few million years) according to a consensus of multiple molecular clock studies, and that exhibit a morphological disparity in their body plans comparable to, say, Pakicetus and Basilosaurus. To be clear, of course no evolutionist ever claimed that Pakicetus was the actual ancestor of Basilosaurus. It rather represented a side branch of the cetacean stem group. But what evolutionists definitely do imply is that the stem species was roughly similar in body plan to Raoellidae and Pakicetidae. Therefore, this challenge is absolutely valid and reasonable.
An obvious possible objection by Darwinists might be that recent species pairs do not represent ancestor-descendent lineages but just cousin lineages that both diverged from a common ancestor. Yes, I get it. However, this also applies for most fossil examples, and there is a catch: While differences in ancestor-descendent lineages could only accumulate in a single evolving lineage, recent lineages could both evolve differences during the same time in each lineage and thus should rather present more and not less morphological disparity. Therefore, this point makes the problem even worse for Darwinists.
Maybe evolutionists will appeal to yet unknown non-Darwinian processes. However, the great advantage of this new argument is that it is totally independent of the nature of the transformation process. You could simply consider that process as a black box. Therefore, it is totally irrelevant if Darwinists invent some new possible mechanism. The crucial point is not the process, but the resulting pattern of new body plans consistently having come into being abruptly in the distant past, but not in the more recent past.
No Conceivable Reason
There is no conceivable reason why a disparity like that between Pakicetus and Basilosaurus should be limited to the fossil record, where it can be found in numerous examples among all groups of organisms, while being totally absent among the millions of recent species. So, let’s be generous and not restrict the challenge to the TimeTree database. Just find any pair of species among the millions of living species to meet the challenge. Only one! Come on, if unguided evolution really can do its magic, this should not be too difficult, should it? Well, I won’t hold my breath, but if the challenge cannot be met, Darwinists should be asked to explain why.
Here is my explanation. Darwinism is wrong, and this applies not only to the neo-Darwinian process of random mutation and natural selection but to any unguided evolutionary processes including those suggested by proponents of the so-called Extended Synthesis (e.g., Shapiro et al. 2014, Laland et al. 2014, 2015, Garte 2016, Müller 2016, 2017).
There is no evolutionary reason why the creative power of this process should have been active over all of Earth history but then ceased to function within the past 10 million years. Intelligent design proponents can easily explain this pattern: there was creative intelligent intervention in the history of life, but this creative activity deliberately ceased with the arrival of humans as the final telos. Any further explanation would have to transgress the methodological limits of the design inference, but Judeo-Christian theists will certainly recognize an eerie correspondence with the Biblical message, which says that God rested from his creative activity after the creation of humans (Genesis 2:2-3).
References
- Alroy J 2002. How many named species are valid? PNAS 99(6), 3706–3711. DOI: 10.1073/pnas.062691099.
- Bechly G 2018. Alleged Refutation of the Cambrian Explosion Confirms Abruptness, Vindicates Meyer.Evolution News May 29, 2018.
- Bechly G 2021. Chapter 31: Does the Fossil Record Demonstrate Darwinian Evolution? Pp 345–356 in: Dembski WA, Luskin C, Holden JM (eds). The Comprehensive Guide to Science and Faith. Eugene (OR): Harvest House.
- Bechly G, Meyer SC 2017. Chapter 10. The Fossil Record and Universal Common Ancestry. Pp 331–361 in: Moreland JP, Meyer SC, Shaw C, Gauger AK, Grudem W (eds). Theistic Evolution: A Scientific, Philosophical, and Theological Critique. Wheaton (IL): Crossway.
- Chapman AD 2009. Numbers of Living Species in Australia and the World. 2nd ed. Canberra (Australia): Australian Biological Resources Study. [PDF]
- Daley AC, Antcliffe JB, Drage HB, Pates S 2018. Early fossil record of Euarthropoda and the Cambrian Explosion. PNAS 115(21), 5323–5331. DOI: 10.1073/pnas.1719962115.
- Garte S 2016. New Ideas in Evolutionary Biology: From NDMS to EES. Perspectives on Science and Christian Faith 68, 3–11. [PDF]
- Hedges SB, Kumar S (eds.) 2009. The Timetree of Life. New York (NY): Oxford University Press.
- Hedges SB, Dudley J, Kumar S 2006. TimeTree: a public knowledge-base of divergence times among organisms. Bioinformatics 22, 2971–2972. DOI: 10.1093/bioinformatics/btl505.
- Hedges SB, Marin J, Suleski M, Paymer M, Kumar S 2015. Tree of Life Reveals Clock-Like Speciation and Diversification. Molecular Biology and Evolution 32, 835–845. DOI: 10.1093/molbev/msv037.
- Helbig AJ, Seibold I, Martens J, Wink M 1995. Genetic Differentiation and Phylogenetic Relationships of Bonelli’s Warbler Phylloscopus bonelli and Green Warbler P. nitidus. Journal of Avian Biology 26(2), 139–153. DOI: 10.2307/3677063.
- IISE 2012. 2011 SOS State of Observed Species. Tempe (AZ): International Institute for Species Exploration. [Website]
- Kimura Y, Hawkins MTR, McDonough MM, Jacobs LL, Flynn LJ 2015. Corrected placement of Mus–Rattus fossil calibration forces precision in the molecular tree of rodents. Scientific Reports 5: 14444, 1–9. DOI: 10.1038/srep14444.
- Kumar S, Stecher G, Suleski M, Hedges SB 2017. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Molecular Biology and Evolution 34, 1812–1819. DOI: 10.1093/molbev/msx116.
- Laland KN, Uller T, Feldman MW, Sterelny K, Müller GM, Moczek A, Jablonka E, Odling-Smee J, Wray GA, Hoekstra HE, Futuyma DJ, Lenski RE, Mackay TFC, Schluter D, Strassmann JE 2014. Does evolutionary theory need a rethink? Nature 514, 162–164. DOI: 10.1038/514161a.
- Laland KN, Uller T, Feldman MW, Sterelny K, Müller GM, Moczek A, Jablonka E, Odling-Smee J 2015. The extended evolutionary synthesis: its structure, assumptions and predictions. Proceedings of the Royal Society B 282:20151019, 1–14. DOI: 10.1098/rspb.2015.1019.
- Latty T, Lee T 2019. How many species on Earth? Why that’s a simple question but hard to answer. The Conversation April 28, 2019.
- Locey KJ, Lennon JT 2016. Scaling laws predict global microbial diversity. PNAS 113(21), 5970–5975. DOI: 10.1073/pnas.1521291113.
- Long Story Short 2020. Ep. 2 Whale Evolution: Good Evidence for Darwin? Discovery Science YouTube channel 24.04.2020.
- May RM 1988. How many species are there on earth? Science 241(4872), 1441–1449. DOI: 10.1126/science.241.4872.1441.
- Mondolfi E, Boede EO 1981. A hybrid of a spectacled bear (Tremarctos ornatus) and an Asiatic black bear (Selenarctos thibetanus) born at the Maracay Zoological Park, Venezuela. Memoria de la Sociedad de Ciencias Naturales La Salle 41(115), 143–148. [ResearchGate]
- Mora C, Tittensor DP, Adl S, Simpson AGB, Worm B 2011. How Many Species Are There on Earth and in the Ocean? PLoS Biology 9(8): e1001127, 1–8. DOI: 10.1371/journal.pbio.1001127.
- Müller G 2016. The extended evolutionary synthesis. Talk presented at the scientific meeting New Trends in Evolutionary Biology at The Royal Society, November 7-9, 2016, London. https://royalsociety.org/science-events-and-lectures/2016/11/evolutionary-biology/
- Müller G 2017. Why an extended evolutionary synthesis is necessary. Interface Focus 7:20170015, 1–11. DOI: 10.1098/rsfs.2017.0015.
- Prothero DR 2014. Species Longevity in North American Fossil Mammals. Integrative Zoology 9, 383–393. DOI: 10.1111/1749-4877.12054.
- Raup DM 1976. Species diversity in the Phanerozoic: a tabulation. Paleobiology 2(4), 279–288. DOI: 10.1017/s0094837300004917.
- Raup DM 1991. Extinction: Bad Genes or Bad Luck? New York (NY): W.W. Norton. [also see New Scientist 13 September 1991]
- Rohland N, Malaspinas A-S, Pollack J, Slatkin M, Matheus P, Hofreiter M 2007. Proboscidean Mitogenomics: Chronology and Mode of Elephant Evolution Using Mastodon as Outgroup. PLOS Biology 5(8): e207, 1663–1671. DOI: 10.1371/journal.pbio.0050207.
- Shapiro JA, Noble D, Pookottil R 2014. The Third Way of Evolution. Website: https://www.thethirdwayofevolution.com.
- Strain D 2011. 8.7 Million: A New Estimate for All The Complex Species on Earth. Science 333, 1083. DOI: 10.1126/science.333.6046.1083.
- Sweetlove L 2011. Number of species on Earth tagged at 8.7 million. Nature 23 August 2011. DOI: 10.1038/news.2011.498.
- Tangley L 1997. How many species are there? US News & World Report 8/10/97.
- Teichert C 1956. How Many Fossil Species? Journal of Paleontology 30(4), 967–969. JSTOR: 1300432.
- Valentine JW 1970. How Many Marine Invertebrate Fossil Species? A New Approximation. Journal of Paleontology 44(3), 410–415. JSTOR: 1302578.
Hail the Lord JEHOVAH: the great king!
◄ Psalm 83 ►
American Standard Version
God, Don't Keep Silent
1O God, keep not thou silence: Hold not thy peace, and be not still, O God.
2For, lo, thine enemies make a tumult; And they that hate thee have lifted up the head.
3Thy take crafty counsel against thy people, And consult together against thy hidden ones.
4They have said, Come, and let us cut them off from being a nation; That the name of Israel may be no more in remembrance.
5For they have consulted together with one consent; Against thee do they make a covenant:
6The tents of Edom and the Ishmaelites; Moab, and the Hagarenes;
7Gebal, and Ammon, and Amalek; Philistia with the inhabitants of Tyre:
8Assyria also is joined with them; They have helped the children of Lot. Selah
9Do thou unto them as unto Midian, As to Sisera, as to Jabin, at the river Kishon;
10Who perished at Endor, Who became as dung for the earth.
11Make their nobles like Oreb and Zeeb; Yea, all their princes like Zebah and Zalmunna;
12Who said, Let us take to ourselves in possession The habitations of God.
13O my God, make them like the whirling dust; As stubble before the wind.
14As the fire that burneth the forest, And as the flame that setteth the mountains on fire,
15So pursue them with thy tempest, And terrify them with thy storm.
16Fill their faces with confusion, That they may seek thy name, O JEHOVAH.
17Let them be put to shame and dismayed for ever; Yea, let them be confounded and perish;
18That they may know that thou alone, whose name is JEHOVAH, Art the Most High over all the earth.
◄ Psalm 8 ►
American Standard Version
1O JEHOVAH, our Lord, How excellent is thy name in all the earth, Who hast set thy glory upon the heavens!
2Out of the mouth of babes and sucklings hast thou established strength, Because of thine adversaries, That thou mightest still the enemy and the avenger.
3When I consider thy heavens, the work of thy fingers, The moon and the stars, which thou hast ordained;
4What is man, that thou art mindful of him? And the son of man, that thou visitest him?
5For thou hast made him but little lower than God, And crownest him with glory and honor.
6Thou makest him to have dominion over the works of thy hands; Thou hast put all things under his feet:
7All sheep and oxen, Yea, and the beasts of the field,
8The birds of the heavens, and the fish of the sea, Whatsoever passeth through the paths of the seas.
9O JEHOVAH, our Lord, How excellent is thy name in all the earth!
Saturday, 23 April 2022
On Russia's "Cosa Nostra"
<iframe width="853" height="480" src="https://www.youtube.com/embed/A6Q72R1aYwA" title="YouTube video player" frameborder="0" allow="accelerometer; autoplay; clipboard-write; encrypted-media; gyroscope; picture-in-picture" allowfullscreen></iframe>
Saving junk DNA?
Dan Graur: The Core Rationale for Claiming “Junk DNA”
See the last entry in this Twitter thread (below), and then our follow-up comments under the screen capture. Dan Graur is a University of Houston biologist and prominent junk DNA advocate. The link to the article in The Scientist that he is grumbling about is included at the very end of this post.
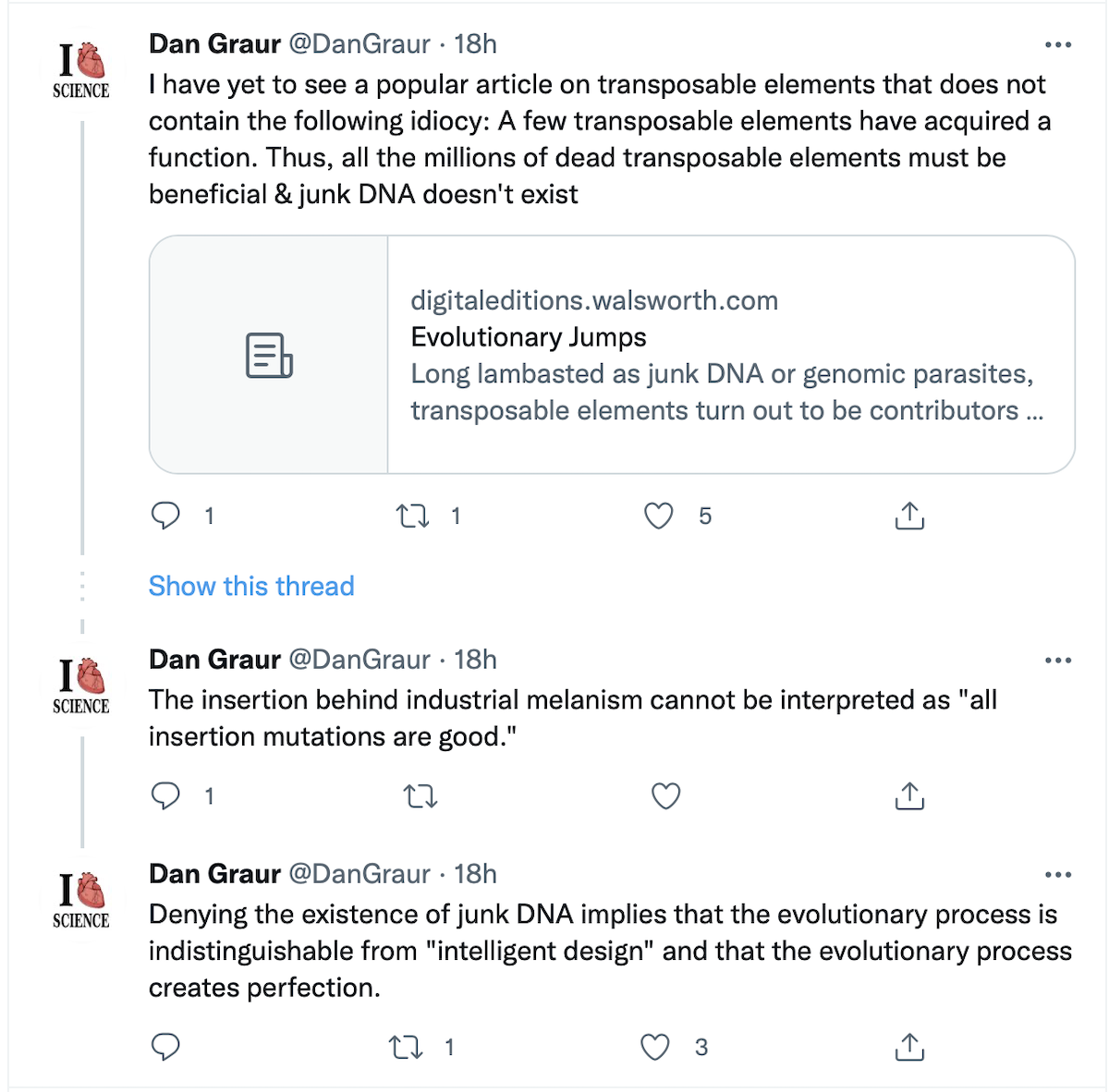
So What’s Wrong with Graur’s Argument?
First, it is unlikely that the growing number of biologists looking for possible functions in so-called “junk DNA” are ID proponents (although a few might be).
Consider this syllogism, however:
- Evolutionary processes lack foresight.
- Processes without foresight create novel functions only infrequently, more often causing non-functionality.
- “Junk DNA” appears to be non-functional.
- Therefore, since evolution is true, it is pointless to look for functions in apparently functionless (“junk”) DNA; what appears to lack function, truly does lack function.
Here’s the Problem
Premises (1) and (2) could be true, as doubtless most biologists would agree, yet (3) does not follow from either premise, and refers in any case only to an appearance, exactly the sort of superficial impression calling for further analysis. Moreover, (4) is a counsel of despair — a GENUINE science-stopper. Since “only infrequently” in premise (2) is defined entirely by the sample size, not by evolutionary theory itself, “no function” claims are empirically unsupportable, resting wholly on negative evidence.
Graur’s argument also affirms the consequent, as tendentious arguments often do. Just take a look at premises (2) and (3) and their logical relation.
That explains why many researchers, who are fully on board with evolution, nonetheless ignore Graur’s advice. They decline to kiss the no-function wall, as Paul Nelson put it back in 2010:
Why Kissing the Wall Is the Worst Possible Heuristic for Biological Discovery
In biology, the claim “structure x has no function” can only topple in one direction, namely, towards the discovery of functions. “No function” represents a brick wall of infinite extent, from which one can only fall backwards, into the waiting arms of a function one didn’t see, or overlooked.
Because one was kissing the wall, so to speak.
Here is the article that got under Graur’s skin: “Evolutionary Jumps,” by Christie Wilcox.
And speaking of election meddling.
<iframe width="784" height="441" src="https://www.youtube.com/embed/f9Q19QJpJ4s" title="YouTube video player" frameborder="0" allow="accelerometer; autoplay; clipboard-write; encrypted-media; gyroscope; picture-in-picture" allowfullscreen></iframe>
Don't give peace a chance?
<iframe width="784" height="441" src="https://www.youtube.com/embed/4IbOUqWbumE" title="YouTube video player" frameborder="0" allow="accelerometer; autoplay; clipboard-write; encrypted-media; gyroscope; picture-in-picture" allowfullscreen></iframe>
Another exposition on engineerless engineering.
Darwinists Seek to Explain the Eye’s Engineering Perfection
Yesterday we looked at a paper by Tom Baden and Dan-Eric Nilsson in Current Biology debunking the old canard that the human eye is a bad design because it is wired backwards. We saw them turn the tables and show that, in terms of performance, the inverted retina is actually as good or better than the everted retina. Vertebrate eyes “come close to perfect,” they said. Ask the eagles with “the most acute vision of any animal,” which would include cephalopods with their allegedly more logical arrangement. Eagles win! Squids lose! Baden and Nilsson looked at eyes from an “engineer’s perspective” and shared good reasons for the inverted arrangement. They even spoke of design seven times; “the inverted retinal design is a blessing,” they argued.
And yet they maintain that eyes evolved by blind, unguided natural processes. How can they believe that? In this follow-up, we look at the strategies they use to maintain the Darwinian narrative despite the evidence.
Personification
First, they turn evolution into an engineer. Personification is a common ploy by Darwinists. Richard Dawkins envisioned a “blind watchmaker” replacing Paley’s artificer. Darwin even gave natural selection a gender:
Natural selection acts only by taking advantage of slight, successive variations. She can never take a great and sudden leap, but must advance by short and sure, though slow steps.
Evolutionists speak freely of natural selection as a “tinkerer” cobbling together whatever odd parts are handy so that a solution, however, awkward, comes to satisfy a need brought on by “selection pressure.” The resulting structures give the illusion of design but are not the work of a rational agent. Neil Thomas calls such stories “agentless acts” by which engineered structures arise by “pure automatism or magical instrumentality quite outside common experience or observability.” Evolutionists need no magician; the rabbit emerges out of the hat spontaneously, as if an invisible hand pulled on its ears. Watch how Baden and Nilsson personify evolution and turn it into a Blind Tweaker and Opportunist:
So, in general, the apparent challenges with an inverted retina seem to have been practically abolished by persistent evolutionary tweaking. In addition, opportunities that come with the inverted retina have been efficiently seized. In terms of performance, vertebrate eyes come close to perfect. [Emphasis added.]
Visualization
A second tactic Baden and Nilsson use is visualization. Figure 3 in their article shows three stages of a possible evolutionary path from a primitive photoreceptor to a “high quality spatial vision” with “usable space” between the lens and retina. “This space could be usefully filled by the addition of neurons that locally pre-process the image picked up by the photoreceptors,” the caption reads. Well, then, what personified tinkerer would not take advantage of such prime real estate? Give the Blind Tweaker room to homestead and you will shortly find him (or her) setting up shop. What’s missing in the visuals are giant leaps over engineered systems in the gaps, and an account of how they all become coordinated to make vision work.
Inevitability
Another tactic used by Baden and Nilsson is the notion of inevitability. Evolution was forced to take the path it did. Evolution had no choice, they imply, because the first time a layer of light-sensitive cells began to invaginate into a cup shape, the path forward was set in concrete.
The specific reason for our own retinal orientation is the way the nervous system was internalised by invagination of the dorsal epidermis into a neural tube in our pre-chordate ancestors. The epithelial orientation in the neural tube is truly inside out. The vertebrate eye cup develops from a frontal (brain) part of the neural tube, where the receptive parts of any sensory cells naturally project inwards into the lumen of the neural tube (the original outside). In contrast, the eyes of octopus and other cephalopods develop from cups formed in the skin, and the original epithelial outside keeps its orientation.
This tactic gives a Darwinist the flexibility to use the same theory to explain opposite things. Evolution is so rigid it must follow the path the ancestor took, but so malleable that all subsequent engineering can be optimized to perfection. Surely, though, if natural selection has the magical powers the Darwinists ascribe to it, it could have ditched ancestral traditions and remodeled the eye cup. Isn’t that how all innovations begin in the theory?
Airy Nothings
The sneakiest tactic used by the authors is what Neil Thomas described as “notional terms.” These are “airy nothings” and “empty signifiers” that gloss over difficulties by replacing evidence with factoids. A factoid, Thomas says, is “a contention without empirical foundation or any locatable referent in the tangible world but one nevertheless held to be true by the person who proposes it” (italics in original). To use this tactic, just assume that your notion is true. Complex things evolved. They originated. They emerged. They arose. They started. They came. They developed. They improved.
Bipolar cells emerged, slotting in between the cones and the ganglion cells to provide a second synaptic layer right in the sensory periphery. Further finesse came through the addition of horizontal and amacrine cells. The result is a structurally highly stereotyped sheet of neuronal tissue present in all extant vertebrates….
Notional terms like this, that assume what need to be proved, can slip through unnoticed unless one is on guard for them. The following contains two of them, while distracting readers with the ID-friendly argument that the eye is not flawed.
Arguments for a basically flawed orientation of the vertebrate retina are built around an eye that we encounter in a grossly enlarged state compared to its humbler origins. We are now more than 500 million years down the road from where vertebrate vision started in the early Cambrian.
When the readers weren’t paying attention, they were being told that the eye had humble origins in the early Cambrian. A truer statement would have read, “Arguments for a basically flawed orientation of the vertebrate retina are built around the assumption of Darwinian evolution.”
Consensus
Baden and Nilsson might counter that they don’t need to give any details about how evolution achieved engineering perfection, because the scientific community has already reached a consensus that Darwinian evolution explains everything in biology, so they can just take it for granted. This is a form of argument from authority, but worse in this instance. As shown in the previous post, the strongest case for eye evolution — so strong that Richard Dawkins celebrated it publicly — was the 1994 graphic that Jonathan Wells and David Berlinski exposed, and Nilsson was one of the perpetrators! To take that for granted would be like drawing a picture of a unicorn in 1994 and then using that drawing as evidence for unicorns in 2022. Evolutionists leaped onto that 1994 article because nothing better had shown up since Darwin got cold shudders considering this “organ of extreme perfection,” the eye.
The Power of Master Narratives
A doctorate in a relatively rare field of expertise called the Rhetoric of Science is held by Thomas Woodward, author of Doubts About Darwin (2003). In that book he examines the role of “fantasy themes” (which he prefers to call “projection themes”) in the history of the Darwin versus Design debate. One of these is the “progress narrative” that the universe is on a continuous path toward higher complexity (p. 52). These projection themes — not empirical data — were of paramount importance in the wide acceptance of evolution when the Victorian atmosphere was fragrant with feelings of progress. Darwin and his successors had a simple, compelling story that was part “factual-empirical narrative” and part “semi-imaginative narrative” (p. 22).
To understand how Darwinians continue to maintain what appears to ID advocates as cognitive dissonance, i.e., that nature is exquisitely engineered but emerged by blind processes, observers need to be cognizant of the projection themes and rhetorical tactics Darwinians use to forestall a Design Revolution. The ones used by Baden and Nilsson cannot be conquered merely by appeals to empirical facts. They need to be exposed as the ploys they are and supplanted by better narratives founded on stronger evidence and logic.
Charles Darwin the design advocate?
Charles Darwin’s “Intelligent Design”
I wrote here yesterday about Charles Darwin’s orchid book. Shortly after its publication, reviews of the book began appearing in the British press. Unlike with the Origin, the reviews were overwhelmingly positive. Reviewers were extremely impressed with Darwin’s detailed documentation of the variety of contrivances in orchids. But much to Darwin’s dismay, they did not see this as evidence of natural selection.
An anonymous reviewer in the Annals and Magazine of Natural History wrote in response to Darwin’s contention that nature abhors perpetual self-fertilization:
Apart from this theory and that of ‘natural selection,’ which we cannot think is much advanced by the present volume, we must welcome this work of Mr. Darwin’s as a most important and interesting addition to botanical literature.
Other reviewers went much further. M. J. Berkeley, writing in the London Review, said:
…the whole series of the Bridgewater Treatises will not afford so striking a set of arguments in favour of natural theology as those which he has here displayed.
Marvels of Divine Handiwork
A review by R. Vaughn in the British Quarterly Review opined:
No one acquainted with even the very rudiments of botany will have any difficulty in understanding the book before us, and no one without such acquaintance need hesitate to commence the study of it. For, in the first place, it is full of the marvels of Divine handiwork.
According to the Saturday Review:
By contrivances so wonderful and manifold, that, after reading Mr. Darwin’s enumeration of them, we felt a certain awe steal over the mind, as in the presence of a new revelation of the mysteriousness of creation.
“New and Marvelous Instances of Design”
Even Darwin’s pigeon-fancier friend, William Tegetmeier, noted the existence in the book of “new and marvelous instances of design.” And an anonymous reviewer in the British and Foreign Medico-Chirurgical Review wrote:
To those whose delight it is to dwell upon the manifold instances of intelligent design which everywhere surround us, this book will be a rich storehouse.
Darwin’s “flank movement on the enemy” failed miserably. Unable to make a convincing case for natural selection in his broader species work, he tried instead to stealthily impress the scientific world by appeal to the exquisite variety of fertilization methods among orchids. Darwin impressed the scientific world alright. He showed how difficult it is to understand the variety of living organisms without appeal to design.
His orchid book may well be the most important of all Darwin’s publications. It made a unique contribution to 19th-century natural history — or is that natural theology? I can think of no greater irony than the fact that Charles Darwin, who Richard Dawkins felt made it possible to be an intellectually fulfilled atheist, actually bequeathed to 19th-century natural historians one of the most impressive cases for intelligent design ever made.
Subscribe to:
Comments (Atom)