At one time, some Bible minimalists (those who believe the Bible is of minimal value historically) questioned the very existence of King David. In recent years, numerous archaeological discoveries have confirmed the existence of Israel’s greatest king, and affirmed numerous details in the biblical text regarding his life and the times in which he lived. Here are the top ten discoveries related to King David.
10. Ancient Slingshots
David is perhaps best-known for his epic mano-a-mano battle against Goliath. The boy with a sling defeated a gigantic, seasoned warrior. While many are familiar with the Y-shaped slingshots with the rubber bands that are used today, slings in the Old Testament were quite different. Biblical weapons expert, Dr. Boyd Seevers describes them: “A sling can be a simple as a strap some three feet in length and one inch in width, though it is often made with two narrow chords attached to a wider pouch in the center. Often, the sling is woven from wool or some other type of flexible material from an animal or plant. One end is looped or knotted to attach to one of the fingers of the thrower’s hand, and the other end is knotted for the thumb and forefinger to grip until the moment of release.”1 Several ancient slingshots from Egypt have survived until today, including King Tut’s sling, which was discovered in his tomb. Slingshots were formidable long-range weapons in antiquity. Ancient texts suggest that slingers were accurate with their projectiles up to 400 yards.2Scripture records that there were 700 men from the tribe of Benjamin who could “sling a stone at a hair and not miss.” (Judges 20:16). This gives us a better understanding of the advantage David had in his battle with Goliath. Of course, we ought not forget that the Battle belonged to the Lord (1 Sam. 17:47).
9. The Gath Ostracon
This inscribed pottery sherd was discovered at Tell es-Safi/Gath and contains two names that are similar etymologically to the name Goliath. Photo: The Tell es-Safi/Gath Archaeological Project
In 2005, an inscribed ostracon (inscribed pottery sherd) was unearthed at Tell es-Safi (the site of the biblical Philistine city of Gath) that was dated to the Iron Age 2A period (when David and Goliath lived). The inscription, written in Semitic “Proto-Canaanite” script contained two names: ALWT and WLT.3 Both of these names (ALWT – Heb. אלות and WLT – Heb. ולת) are very similar etymologically to the name Goliath (Heb. גלית). Aren Maeir, the director of the excavations at Tell es-Safi/Gath summaries the importance of this inscription: “1) the inscription demonstrates that ca. the 10th/9th cent. BCE, names very similar to Goliath were in use at Philistine Gath. This does provide some cultural background for the David/Goliath story; 2) that already early in Iron IIA, the Philistines adopted the Semitic writing systems.”4
8. Hebron (Tell Hebron/Tell
David initially reigned as king of Judah at Hebron, while Saul’s son Ish-Bosheth, reigned as king of Israel from Mahanaim. (2 Sam. 2:8-11). Hebron has identified as Jebel er-Rumeide, also called Tell Rumeide or Tell Hebron. Five LMLK (to the king) jar handles bearing the city name Hebron have been discovered at the site.5 Excavations have uncovered sections of the Middle Bronze II city wall, which continued to be used in the Iron I and II periods,6 as well as the remains of four-roomed houses and fragments of collared-rim jars, both of which are typically associated with Israelites.7 The remains of David’s royal residence likely lie on the summit of the tell, which is covered by a medieval structure (called Deir Arbain by the locals) which was originally a Byzantine monastery, and is off-limits to excavation.8 While David is more commonly associated with Jerusalem, the first capital of the Kingdom of Judah was at Hebron.
7. Geshur (et-Tell)
While David was reigning in Hebron, he had numerous sons by various wives. His thirdborn was Absalom the son of Maacah daughter of Talmai king of Geshur (2 Sam. 3:3). Later, after Absalom had murdered his brother Amnon, he fled to his grandfather, Talmai, son of Ammihud, the king of Geshur, and lived with him for three years (2 Sam. 13:37-38). Scholars have suggested that David married Maacah, the daughter of the king of Geshur to solidify relations between their two kingdoms, and to strengthen his own position. In antiquity, the usual practice was for the daughter of the more powerful ruler to be given to the weaker ruler, which would indicate that Geshur was the stronger kingdom.9
Et-Tell, a site 3km (1.5 miles) from the north-east shore of the Sea of Galilee has been identified as the capital of the Kingdom of Geshur. It satisfies the geographic criteria in Scripture (Deut 3:14; Josh 12:4–5; 13: 11–13), where it often paired with Abel Beth Maacah. Abel Beth Maacah is identified as Tell Abil al-Qamh, and et-Tell is identified as the capital of the kingdom of Geshur; both sites are the most prominent Iron Age mounds in the region.10 Et-Tell (one of the contending sites for New Testament Bethsaida), was a significant fortified city in David’s day; the massive four–chamber Iron-Age Gate can still be seen today. A carved basalt stone stela was discovered near the city gates and depicts a bull-headed figure, which likely represents either the storm god or the moon god the people of Geshur worshiped.11 If the identification of et-Tell as the capital of the kingdom of Geshur is correct, then this is likely where David’s wife Macaah was from, and the place his son Absalom lived for three years.
6. Large Stone Structure (King David’s Palace in Jerusalem)
David was eventually made king over all Israel (2 Sam. 5:3), and he immediately captured Jerusalem (2 Sam. 5:7), and set to budling a palace, which Hiram, king of Tyre, assisted in the construction of by providing cedar logs, carpenters, and stonemasons (2 Sam. 5:11). From 2005-2007, Israeli archaeologist, Dr. Eilat Mazar, unearthed what is now known as the Large Stone Structure, a monumental building complex with walls that were 6-8 feet wide, constructed of impressive stones, and to which a beautiful 5-foot-long proto-Aeolic capital likely once belonged. It is located above the famous Iron-Age Stepped-Stone Structure, which probably supported the Fortress of Zion and the Large Stone Structure above. The pottery found beneath the Large Stone Structure, dated the first phase of its construction to the beginning of the Iron Age IIa (10th century BC), the time of King David. Based on the palatial nature of the structure and the fact that its location matched biblical data (such as 2 Sam. 5:17 – David descending from his residence to the fortress), Mazar identified the structure as David’s Palace.12 While this identification has not been without controversy, numerous scholars accept her conclusion. Archaeologist, Dr. Scott Stripling, states, “Eilat Mazar’s excavation of the Large Stone Structure likely revealed David’s actual palace, just above the well-known Stepped Stone Structure or milo.”13 Nadav Na’aman, former professor of Jewish History at Tel Aviv University notes, “The Large Stone Structure, which Eilat Mazar unearthed and identified as the residence of King David, is indeed a suitable candidate for this building, or more accurately, for its northeastern wing.”14
5. Judahite Cities (Khirbet Qieyafa and Tel Eton)
Scripture records David’s kingdom expanding (2 Sam. 8:1-4), and controlling the kingdom from his capital city of Jerusalem. Two fortified sites dating to the 10th century have been unearthed which scholars believe are evidence of such a centralized authority Yosef Garfinkel (Hebrew University) and Saar Ganor (Israel Antiquities Authority) oversaw excavations at Khirbet Qeiyafa from 2007-2013. The site is located 30km southwest of Jerusalem, within the kingdom of Judah, and surrounded by a massive casement fortification wall with two gates. Its 10th century date was confirmed by radiocarbon dating of pits in a destruction layer of a large royal storeroom.15 The excavators identified it as a Judahite outpost based on inscriptional evidence (an ostracon with one of the earliest Hebrew texts yet found), a lack of pig bones, and the presence of cultic shrines that did not have any graven images of people or animals. Garfinkel and Ganor state, “The massive construction of the Khirbet Qeiyafa city wall, which required 200,000 tons of stone, and the massive eastern gate of the city with two stones of ca. 10 tons each, proclaim the power and authority of a centralize political organization, namely a state.”16controlling the Archaeologists from Bar-Ilan University recently excavated El Eton, another site that dates to the time of David and displays evidence of a strong, central political administration during its construction. A monumental structure, dubbed the “governor’s residency” was built using quality ashlar stones in the typical Israelite four-room design. Radiocarbon dating of samples taken from the foundation deposit indicate that the earliest phase of the structure was built in the late 11th-10th century BC. In an article in the journal Radiocarbon, Avraham Faust and Yair Sapir wrote: “The building of the ‘governor’s residency,’…suggests that the settlement at Tel ‘Eton was transformed in the 10th century BCE, lending important support to the historicity of the United Monarchy…[it] exhibits the earliest evidence for the use of ashlar stones in the region of Judah, and the mere erection of this edifice challenges one of the arguments against the historical plausibility of the United Monarchy (i.e., that ashlar construction appeared hundreds of years later).”17
4. Davidic Kings
After King David’s death, 20 of his descendants ruled in succession after him, from Solomon to the kings who reigned over the southern Kingdom of Judah. Archaeology has furnished numerous finds attesting to many of these Davidic kings.18 The nearly identical gates at Hazor, Megiddo, and Gezer are evidence of Solomon’s building activity as described in 1 Kings 9:15.19 Ahaziah is the king of the “house of David” referred to on the Tel Dan Stele (see below). Two seals which once belonged to officials in King Uzziah’s court mention him by name. A bulla (clay seal impressions) that reads, “Belonging to Ahaz (son of) Yehotam [Jotham], King of Judah” is held in the private collection of Shlomo Moussaieff in London. Numerous seal impressions from King Hezekiah have been discovered, and he is named in the annals of Sennacherib. Manasseh is named in the annals of both Esarhaddon and Ashurbanipal, while Jehoiachin is mentioned in ration tablets from Babylon. Each of these discoveries independently corroborates the biblical description of a Davidic line of kings who reigned in Israel and Judah for generations.
3. “Heights of David” Inscription
King David’s name has been found in numerous ancient Inscriptions, including one possible reference from Egypt. When the Egyptian Pharaoh Shishak (Shoshenq I), returned from his campaign in Palestine in 926/25 BC, he commanded that his victories be recorded on the walls of the Temple of Amun in Karnak. More than 150 hieroglyphic name-rings, each represented as a bound prisoner, are recorded on Bubastite Portal detailing the places he conquered during his northern campaign. Names rings 105 and 106 together read h(y)dbt dwt – the “Heights or Highlands of Dawit.” Egyptologist, Kenneth Kitchen, has proposed that this should read, “Heights of David.” He writes, “In Egyptian transcriptions of foreign names (both places and personal), a t could and sometimes did transcribe a Semitic d. This happens in the New Kingdom in such familiar place-names as Megiddo (Egyp. Mkt).”20 He further points to a sixth century Ethiopic inscription citing Psalm 65:19 from the “Psalms of Dawit,” the exact consonants on the Shishak Inscription. Kitchen summarizes: “This would give us a place name that commemorated David in the Negev barely fifty years after his death, within living memory of the man.”21 His conclusion is not without its critics, however, as some have suggested that ring 106 is difficult to decipher and may not read Dawit at all, let alone reference David.
2. Mesha Stele (Moabite Stone)
The famous Meshe Stele (Moabite Stone) consists of 57 fragments which were purchased from bedouin in the 19th century, and assembled, along with a squeeze of the inscription which had been taken before the monument was broken. The stele, a black basalt monument measuring 1.5m (45.28”) high by 60-80cm (23.6-31.5”) wide, is a victory stele of Mesha, king of Moab. Written in Moabite, it describes the same events in 2 Kings 3, Moab’s rebellion against Israelite subjection. In 1994, epigrapher, André Lemaire announced that he had detected a previously-overlooked letter, resulting in the phrase, “House of David.” He wrote: My own examination of the stone and the squeeze, which is now being restored and cleaned of accumulated dust, confirms that t follows the b. I would now, for the first time, reconstruct the missing letter as a d (d). The result: bt[d]wd (dw[d]tb), the “House of [D]avid!”22 The relavent part of the inscription reads, “And the house[of Da]vid dwelt in Horanaim […] and Chemosh said to me: ‘Go down! Fight against Horanaim.’ And I went down, and [I fought against the town, and I took it and] Chemosh [resto]red it in my days” (lines 31-33).23 In 2019, the Mesha Stele hit the news again when Israel Finkelstein, Nadav Na’aman, and Thomas Römer published a paper in the Journal of the Institute of Archaeology of Tel Aviv University analyzeing Line 31 on the Moabite Stone arguing the words in question refer to Balak, not the “House of David.”24 Scholar, Michael Langlois, responded with an analysis published in the Journal Semitica, which high resolution images and Polynomial Texture Mapping (PTM) of the stele to create a 3-D image. The new technology revealed a previously overlooked dot, indicating a break between words, which comes exactly after the area interpreted “House of David,” confirming Lemaire’s initial reading.25
For many patients, blood transfusion is not an option, whether for religious reasons or safety concerns. The Center for Bloodless Medicine and Surgery team understands and respects this view. We are experts in patient blood management, improving health outcomes and creating peace of mind for those who seek the very best healthcare, without the use of blood or blood products.
Discover the Benefits
There are many benefits to a bloodless approach. Research shows that patients who do not receive blood transfusions recover faster, experience fewer infections and leave the hospital sooner than those who do.How it Works: Bloodless Medicine and Surgery - An Alternative to Blood Transfusion
Bloodless medicine and surgery is an alternative to blood transfusion that among other benefits, has been shown to reduce infections and help patients recover faster. In this video, experts from Johns Hopkins explain the techniques used before, during and after surgery to help patients minimize blood loss and the need to receive donated blood.Bloodless Medicine | Hazel's Story
Hazel Skinner has suffered from chronic illnesses for over 15 years and came to Johns Hopkins from Pennsylvania. Hazel's sister, Dawn, has been to every appointment with her and finds comfort in knowing Hazel's care is a top priority for the Bloodless medicine team. Hazel was admitted to Hopkins and was soon introduced to Dr. Linda Resar, the consultant hematologist for the Bloodless program. Hazel recalls researching the Bloodless program after being told she needed to increase her blood count in able to continue with dialysis. While being sensitive to Hazel’s conscientious refusal to accept blood products, Dr. Resar was able to give her the care she needed to get her hemoglobin back up to healthy levels safely without blood transfusions. After discharge, the Bloodless team continues to follow Hazel. Nurse Coordinator Liz Dackiw calls Hazel regularly to make sure her hemoglobin remains at a healthy level and Hazel continues to be seen by the Bloodless team for regular check-ups throughout the year.
Michael Behe’s introduction of irreducibly complex (IC) molecular machines in Darwin’s Black Box is a gift that keeps on giving. Many readers probably had never heard of cilia or flagella back in 1996. The fact that those machines still make useful illustrations of IC now, even more powerfully than they did 25 years ago, is a strong affirmation of his thesis that IC gives evidence of intelligent design. The bacterial flagellum tends to get more mentions because it is such a cool outboard motor that laypersons can immediately relate to. No less wondrous, though a little more obscure, is the cilium.
Behe updated his description the cilium in his second book, The Edge of Evolution (2007), but research has continued apace. The cilium nails the case for intelligent design more than ever, especially when considering how the organelle is built. Inside those tiny hairlike projections is an advanced transportation system that looks for all the world like a motorized two-way railcar inside a mine shaft!
In Current Biology, Gaia Pigino wrote a “Primer” on Intraflagellar Transport (IFT). It’s called intraflagellar because a cilium is a type of flagellum (Latin for whip), which in the generic sense means a whiplike structure that can move. Both cilia and flagella use the IFT system for construction because both need to transport their building blocks down a shaft from the base to the distal tip. From the railcar’s perspective, the tip would seem a long way away.
There are motile cilia, like the ones that keep our windpipes clean and propel sperm cells, and “primary” cilia, which act as sensory antennae on almost all cells. Accurate construction of cilia is vital. When things go wrong, a host of problems called ciliopathies can result in severe diseases and death. Evolution News has mentioned these briefly in previous years (here, here, and here).
Parts List
Consider first how many players are needed to build a cilium. Pigino’s parts list begins with microtubules in a 9+2 arrangement going up the cilium from base to tip. The two center microtubules are singlets; the outer ring of 9 are in doublet pairs. Riding on those rails are two engines: kinesin-2, which travels from base to tip (anterograde), and dynein-2, which goes from tip back to the base. Kinesin-2 has a head, stalk, hinge and two “feet” (called heads) that walk on the microtubule while carrying a load; the engine contains six protein subunits. Dynein-2 also has a motor, stalk, linker and tail, and is powered by two AAA+ domains that spend ATP for power. Those are the two engine types, and they work in teams along the microtubules.
IFT proteins are numbered, such as IFT8 and IFT176. IFT complexes, such as IFT-A, is composed of six IFT proteins. IFT-B, with 16 IFT proteins is another complex. These ride along the trains to the tip, acting as adaptors for the cargo, which include tubulin proteins, dynein parts, membrane proteins and other IFT proteins.
At the base, a basal body structure called the BBSome forms out of eight BB proteins. It functions as the cargo adaptor for the anterograde train. It authenticates other molecules trying to enter the cilium and moves cargo exiting the retrograde train. Overall, “About 24 different proteins constitute the theoretical minimal functional unit of IFT,” Pigino says, although much needs to be learned.
To address the many fascinating questions that remain about the function and mechanisms of IFT within the cilium and beyond will require the development of new technologies. Fortunately, recent years have seen the introduction of approaches such as cryo-FIB scanning electron microscopy, cryo-correlative light and electron microscopy, and expansion microscopy. The opportunity to combine such approaches with in situ protein tagging for EM, isolation of active IFT complexes, and in vitro reconstitution and reactivation of IFT machinery suggests that IFT studies will continue to yield important insights long into the future. [Emphasis added.]
A precise sequence of amino acids is required for each protein’s function, and the longer the protein, the more improbable that chance could get it right. IFT proteins are large. For instance, BBS1 in the BBSome has 593 amino acid residues; IFT172 (part of the IFT-B complex) has 1,749. The improbability is exacerbated when proteins have to work together. It’s not necessary to belabor the point again, but it’s instructive that Pigino never mentions evolution in her article.
Riding the Train
Moving cargo up and down the cilium takes place in five steps. First, the train assembles at the base. Kinesins line up along a microtubule doublet, their “heads” touching the tracks. Parts of dynein (the return engine) are loaded so as not to touch the tracks, avoiding a “tug-of-war” between the engines. Membrane parts and other cargoes are loaded with the help of IFT-A and IFT-B. Like a well-organized monorail car, the completed train “walks” up the track aided by multiple kinesin-2 engines powered by ATP.
At the tip, the third phase begins. Cargo is unloaded and ferried to the growing cilium (microtubules and membrane). Concurrently, the dynein engines are assembled in an “open configuration as an intermediate state to ensure a controlled activation.” The kinesins are disassembled for transport back to the base. The fourth stage activates the dyneins and starts the train moving, carrying both IFT complexes and waste products to the base. The fifth and final stage unloads the cargo, disassembles the retrograde train and recycles the parts. If you conceive of railcars in a narrow mine shaft carrying tools needed by miners at the far end, and returning the carts with waste products, the analogy seems apt — only the cell’s actions are all automated.
It All Has to Work
Pigino spends much of her Primer discussing ciliopathies: the diseases of broken cilia. When the IFT or engine parts have mutations, or the cilium fails to develop properly, terrible things happen — really terrible things. That’s if the organism (or person) survives at all. Many ciliopathies are not witnessed because the defect causes “major issues during early embryonic development that lead to neonatal death in vertebrates.” She lists 14 known ciliopathies that cause named syndromes, like Bardet-Biedl Syndrome, which causes “rod/cone dystrophy, polydactyly, central obesity, hypogonadism, and kidney dysfunction,” or Retinitis Pigmentosa, which causes blindness. Without getting into the gory details, these ciliar defects can harm the skeleton, eyes, kidneys, brain, or multiple systems in the body at once.
Usually, It Does Work
For the majority of people born with working cilia, here is what they do for us:
Cells need to be able to sense different types of signals, such as chemical and mechanical stimuli, from the extracellular environment in order to properly function. Most eukaryotic cells sense these signals in part through a specialized hair-like organelle, the cilium, that extends from the cell body as a sort of antenna. The signaling and sensory functions of cilia are fundamental during the early stages of embryonic development, when cilia coordinate the establishment of the internal left–right asymmetry that is typical of the vertebrate body. Later, cilia continue to be required for the correct development and function of specific tissues and organs, such as the brain, heart, kidney, liver, and pancreas. Sensory cilia allow us to sense the environment that surrounds us; for instance, we see as a result of the connecting cilia of photoreceptors in our retina, we smell through the sensory cilia at the tips of our olfactory neurons, and we hear thanks to the kinocilia of our sensory hair cells. Motile cilia, which themselves have sensory functions, also work as propeller-like extensions that allow us to breathe because they keep our lungs clean, to reproduce because they propel sperm cells, and even to properly reason because they contribute to the flow of cerebrospinal fluid in our brain ventricles…. Thus, the proper function of cilia is fundamental for human health.
Professor Behe did a wonderful service in introducing these marvelous machines to a wider audience. In 1996, he introduced cilia as examples of irreducible complexity. In 2007, with a decade of new knowledge to draw from, he called cilia examples of “irreducible complexity squared.” Pigina’s Primer on cilia cannot argue with that. If you can breathe, eat, smell, taste, hear, and walk, thank the intelligent designer of cilia that makes these pleasures possible.
Hemoglobin is well known as the molecule that transfers oxygen in blood, but its precursor, heme, is lesser known. Heme is a complex molecule that looks geometrically square, with a single iron atom at the center. The heme family of metalloproteins is responsible for multiple functions in the cell and in the bodies of multicellular organisms, including humans. Our lives depend on heme. When not properly handled, though, it can be dangerous.
What Heme Does
Seven scientists (Galvin Leung et al.) from two UK universities (Leicester, Bristol) explain the significance of heme in their paper, “Unravelling the mechanisms controlling heme supply and demand,” published in PNAS. Their homage to heme is unrestrained, as is their appreciation for how the cell handles this toxic molecule.
Heme is essential for the survival of virtually all living systems and is involved in many fundamental biological processes. It is also implicated as a signaling/regulatory molecule and must be mobilized in response to cellular demands. This presents a complex logistical problem: heme cannot simply diffuse around cells because it is both insoluble and cytotoxic. We show that the cell exhibits exquisite control over release of heme by limiting its availability to one molecule or less within cellular compartments. [Emphasis added.]
Such a description should make a Darwinist shudder. How could such an “exquisite control” system evolve piecemeal? Consider just the making of heme:
Heme is essential for the survival of virtually all living systems — from bacteria, fungi, and yeast, through plants to animals. The family of heme proteins is vast, and heme proteins are responsible for a multitude of functions that are essential for the survival of the cell. To meet the needs of supply and demand for heme in cells, most organisms need to synthesize it. Biosynthesis of the heme cofactor is, therefore, one of the most important metabolic processes in biology; it occurs as an eight-step enzymatic pathway, the last three steps of which occur in the mitochondria.
It takes eight steps to synthesize one heme molecule, and virtually all life needs it — even bacteria, among the simplest of organisms. The other enzymes that construct heme had to already exist before heme could do its job. This is a serious chicken-and-egg problem for the origin of life.
Heme Synthesis
A taste of the complexity of heme synthesis can be had in “Biochemistry, Heme Synthesis,” by Ogun, Joy, and Valentine.
Heme biosynthesis starts in mitochondria with the condensation of succinyl Co-A from the citric acid cycle and an amino acid glycine. They combine to produce a key heme intermediate, 5′-aminolevulinic acid (ALA) in mitochondria catalyzed by the pyridoxal phosphate-requiring (vitamin B6) enzyme, aminolevulinic acid synthase(ALAS). This reaction is the rate-limiting step in the pathway….
That’s just for starters. Those interested in the remaining steps involved in heme synthesis can read four more paragraphs of details like these at the link above. Intermediate forms of the molecule shuttle in and out of a mitochondrion, where special gates control traffic in and out. Multiple other molecules and enzymes, including one metalloprotein containing zinc, are involved in the process.
How could the first cell, by chance, hit on this sequence of steps that would challenge a chemistry grad student? A protocell would have needed to come up with this chemical pathway just to get heme, let alone know what to do with it once it had it. Whoops; it’s toxic, too. How many protocell tryouts died from this essential yet cytotoxic substance before figuring out that heme must be handled with care? Darwinism is dead already — but there’s more.
Heme Supply
The focus of the paper by Leung et al. in PNAS is on how cells distribute heme where it is needed without dying from it.
We suggest an exchange mechanism between protein partners to control supply and demand. Such a mechanism would provide an in-built buffering capacity for heme, enable cells to hoard supplies of heme, and protect the cell against the undesirable effects of heme.
How about that; cells know the law of supply and demand. Where did they learn that? In protocell economics class? They also know how to “hoard supplies” of heme (actually, how to maintain emergency stockpiles). In the recent pandemic, some government officials were aghast to find that emergency stockpiles of PPE (personal protective equipment), required by law, had been raided or not maintained. It led to serious shortages and drastic efforts to refill stockpiles, while patients were dying and healthcare workers were exposed to the virus without protection. Cells do not make such mistakes.
Heme Distribution
Scientists have known all about heme and its functions for decades, but few have investigated how cells distribute it where needed. This is important to know, Leung et al. explain, because “Deficiencies or excesses in cellular heme concentration also have widespread implications in health and disease” such as aging, cardiovascular disease, inflammation, and immune response. Accordingly, “there is a need to understand the logistics of heme supply and demand.”
A cell cannot maintain a “pool” of heme to draw from, as once thought, because heme is a “nuisance” to cells. It tends form free radicals, which are dangerous, and though hydrophobic, it dimerizes in solution, making it unsuitable for delivery to proteins that only need one heme molecule per binding site.
A free molecule of heme can therefore only exist transiently, and if a large reserve of heme is present, the heme molecules would presumably need to be exchanged rapidly between binding partners to remain solubilized, in the same way that heme is solubilized within the interior of other well-known heme proteins (e.g., hemoglobin).
The team constructed a molecular heme sensor that glows when bound to heme. In this way, they could watch the “exquisite control” system in action.
A longstanding question has been to establish the mechanisms that control the supply and demand for cellular heme. … we have developed a heme sensor … that can respond to heme availability…. The results demonstrate that concentrations are typically limited to one molecule or less within cellular compartments. These miniscule amounts of free heme are consistent with a system that sequesters the heme and is able to buffer changes in heme availability while retaining the capability to mobilize heme when and where it is needed. … This exquisite control, in which heme is made available for transfer one molecule at a time, protects the cellagainst the toxic effect of excess heme and offers a simple mechanism for heme-dependent regulation in single-molecule steps.
In effect, the cell maintains “an exchangeable (buffered) heme reservoir” that solves the availability problem while simultaneously protecting the cell from heme’s toxic effects. Free heme (the risky kind) was detected only in “a minute fraction of the entire amount of heme present in the cell” and were most likely short-lived in the process of binding to proteins.
Our experiments are thus consistent with the idea that there is a population of the total heme complement that is bound more weakly and therefore reversibly to heme-binding partner proteins or to other molecules (which might include free amino acids) that can buffer against changes in the heme concentration. … These heme molecules that are weakly bound to buffer molecules, along with the miniscule population of free heme, would constitute a body of exchangeable heme in the cell.
In their model, the buffered heme, attached to its partner, passes quickly to the enzyme or protein needing it, something like a quick pass of the ball from one player to another in basketball or football. In the cell’s game, though, there are millions of balls with millions of players passing the heme balls to the players who need it. Because the free energy of the acceptor is at a lower level, the heme is readily transferred to the acceptor, leaving the partner ready to pick up another heme. At any given time, the cell can be aware of the concentration of available heme by sensing the concentration of heme-binding partners, and supply more as the demand increases.
This exquisite control also provides a mechanism for heme-dependent signaling and regulation, as heme can be supplied discretely, leading to the switching on of proteins in single-molecule steps.
If Darwinism had been essential to their work, they surely would have mentioned it. Instead, they found a mechanism that appears (gasp! Can they say this?) designed for a purpose —
We see clear advantages of such an exchange mechanism between protein partners, designed for the purpose of managing heme supply and demand.
Overkill
To nail the case for design, consider the level of exquisite control in the next hierarchical level up. The human body makes around 250 billion red blood cells per day, and each RBC contains 270 million hemoglobin molecules, each constructed with 4 heme groups. That multiplies out to 27 billion trillion hemes per day!
It’s amazing enough that each cell in the body orchestrates its synthesis and availability of heme. On top of that, the whole body, too, regulates the number of hemoglobin molecules and red blood cells that carry another cytotoxic substance — oxygen — from our lungs to each cell in a safe, regulated, exquisitely controlled manner. Every red-blooded person should take this to heart: we would be walking packages of explosives if it were not for mechanisms “designed for the purpose” of using energy safely for life, love, and transcendent meaning.
As I explained in a post yesterday, a TEDx talk from January of this year by MIT bioengineer Erika DeBenedictis, “It’s Time for Intelligent Design,” argues that biology lacks any “intelligent design,” and thus living systems are “imperfect” with “gigantic mistakes,” and we should “play God” in order to “make biology better.” So what are some flaws in biology that we can make “better”? According to Dr. DeBenedictis, “one of the big limitations of biology are the basic building blocks themselves.” Emily Reeves has already addressed Dr. DeBenedictis’s comments about the optimality of amino acids in the genetic code, noting that the set of amino acids used by life is highly optimal. But DeBenedictis goes further and says that the very makeup of the building blocks of life need a major redesign.
“Kind of Boring”
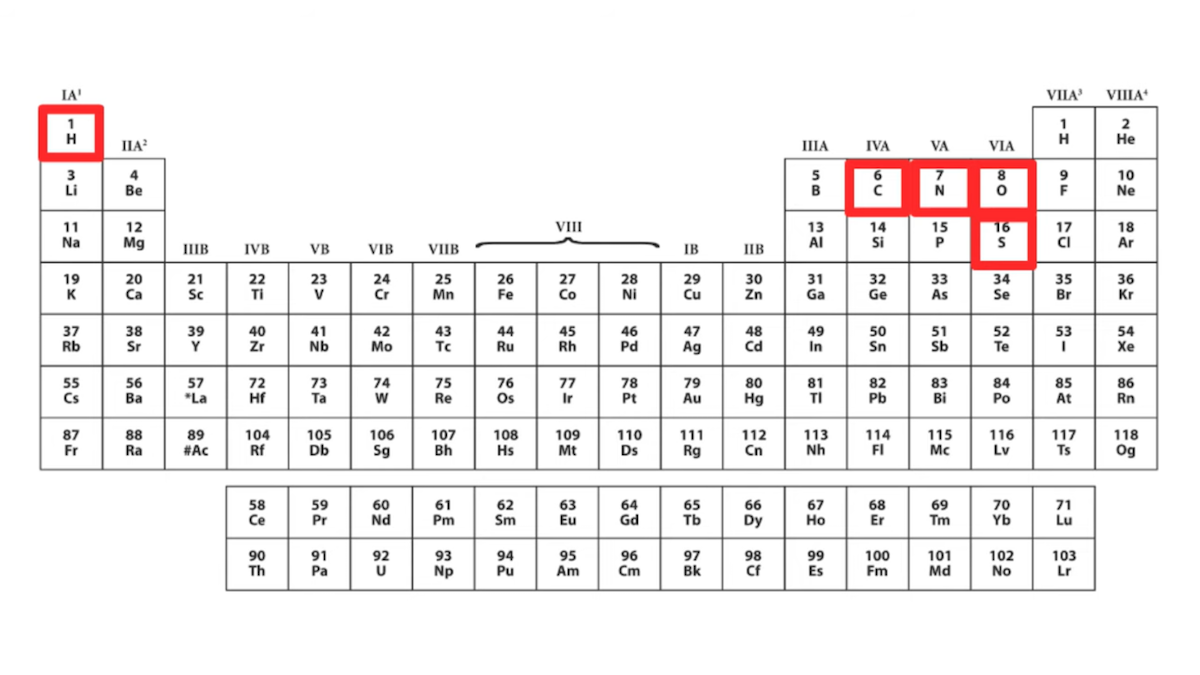
“If we look at the amino acids,” she states, “chemically speaking, they’re actually kind of boring” and “one very easy way to see this is to look at them on a periodic table.” She then shows a periodic table that highlights five chemical elements — hydrogen, carbon, nitrogen, oxygen, and sulfur — she says “Out of all this good stuff, only five chemical elements appear in proteins.” Here’s the slide from the talk:
If we are to take Dr. DeBenedictis at her word, then “only five chemical elements appear in proteins,” namely those highlighted in her chart: hydrogen, carbon, nitrogen, oxygen, and sulfur. She further says she wants to create new building blocks and says, “maybe we could get some heavy metals up here.” The message is that proteins only use five elements and that proteins and enzymes essentially ignore the rest of the period table — more evidence of what she calls “imperfect design.”
Dr. DeBenedictis is right that amino acids themselves don’t contain metals. But is it correct for her to state that “only five chemical elements appear in proteins,” suggesting that the rest of the periodic table is absent from protein complexes and not involved in enzyme chemistry? Is it true that proteins don’t make use of metals? No, that is not correct.
Metals, Metals, Everywhere…
Enzymes are proteins that catalyze reactions, and Voet and Voet’s standard textbook Biochemistry states “Nearly one-third of all known enzymes require the presence of metal ions for catalytic activity.” (Voet and Voet 2004, p. 501). The textbook continues:
There are two classes of metal ion-requiring enzymes that are distinguished by the strengths of their ion-protein interactions:
1. Metalloenzymes contain tightly bound metal ions, most commonly transition metal ions such as Fe2+, Fe3+, Cu2+, Zn2+, Mn2+, or Co3+.
2. Metal-activated enzymes loosely bind metal ions from solution, usually the alkali and alkaline earth metal ions Na+, K+, Mg2+, or Ca2+.
Metal ions participate in the catalytic process in three major ways:
1. By binding to substrates so as to orient them properly for reaction.
2. By mediating oxidation-reduction reactions through reversible changes in the metal ion’s oxidation state.
3. By electrostatically stabilizing or shielding negative charges.
Metal ions are constituents of many metalloproteins, in which they have either catalytic (metalloenzymes) or structural functions. … Metals are important for the biological activity of proteins and their removal or the replacement of one metal by another is often accompanied by a loss of or reduction in the biological activity of the protein. Metal ions in proteins can act as structure promoters or can take part in enzymatic reactions. Divalent metal cations such as Zn2+, Mg2+, Cu2+ and Ca2+ are often associated with the catalytic or regulatory activities of proteins that constitute some of the fundamental chemical life processes. For example, Mg in chlorophyll is important for photosynthesis, Cu (together with Fe) has a role in oxygen-carrying proteins and Zn, Mg and Ca can serve as Lewis acids. (emphasis added, internal citations omitted)
A 2011 entry on metalloenzymes in Springer’s Encyclopedia of Geobiology likewise explains that up to a third of known enzymes — i.e., an important type of protein that fosters chemical reactions in cells — use metals as cofactors:
Metalloenzymes are enzyme proteins containing metal ions (metal cofactors), which are directly bound to the protein or to enzyme-bound nonprotein components (prosthetic groups). About one-third of all enzymes known so far are metalloenzymes… metalloenzymes are found in all enzymes families…
The encyclopedia entry goes on to explain:
Besides enzymes, other metalloproteins are involved in non-enzyme electron transfer reactions (cytochromes), may act as storage (e.g., ferritin for iron) or transport proteins (e.g., transferrin for iron). In the latter groups of proteins, the metal storage is reversible and the metal is a temporary component. Also, ribozymes, i.e., RNA molecules with enzyme function may contain structurally and/or functionally important metal ions … and may be therefore termed as metalloenzymes in a broader sense.
Later the entry lists a table summarizing the functions of various metals in enzymes — here’s a condensed version of that table:
Metallic Element
Roles Played in Enzymes
Potassium
Protein stabilization
Calcium
Protein stabilization
Magnesium
Protein stabilization
Iron
Oxygen transport, storage and/or activation; electron transport; superoxide breakdown
Manganese
Oxygen release; peroxide and superoxide breakdown
Zinc
Protein stabilization; hydrolytic cleavage
Copper
Oxygen transport, storage and/or activation; electron transport; superoxide breakdown
Molybdenum
Oxygen transfer; N2 activation
Cobalt
Free radical reactions; nucleophile
Going through the entry, it becomes evident that the variety and number of uses of metals in enzymes and proteins are almost endless. Examples listed include:
“Mononuclear iron proteins are oxidoreductases (e.g., aromatic amino acid hydroxylases, aromatic ring cleavage dioxygenases, Fe-superoxide dismutase, lipoxygenases, nitrile hydratase, and Rieske oxygenases), or involved in electron transfer (desulforedoxins, rubredoxins, andphotosynthetic reaction centers).”
“Hemoproteins are characterized by an iron porphyrin as prosthetic group” and include “oxidoreductases (catalases, peroxidases), electron transfer proteins (cytochromes), or oxygen transport and storage proteins (globins).”
“Iron–sulfur proteins are characterized by the presence of iron-sulfur clusters containing sulfide-linked di-, tri-, and tetra iron centers in variable oxidation states” and include “ferredoxins, NADH hydrogenases, dehydrogenases, cytochrome creductases, nitrogenases, and other proteins.”
“Copper proteins are involved in oxygen transport or activation processes and electron transport…” and include “ascorbate oxidase, ceruloplasmin, laccase, nitritereductase, auracyanin, and azurin …”
Heavy Metals
Dr. DeBenedictis’s precise statement in her talk says that we should add “heavy metals” to life. It’s not exactly clear how she would define “heavy metals,” but her slide (see above) and language seem to suggest that proteins don’t use any metals at all. While she is likely trying to simplify things to make a point, it is incorrect to say that proteins don’t use heavy metals. For example, a 2020 paper in Scientific Reports states that many “heavy metals” are “bioavailable,” and these include zinc (Zn), copper (Cu), and nickel (Ni). Multiple sources explain how widely used these heavy metals are in plants.
Zinc: A 2016 study notes how important zinc is in plants:
Zinc (Zn) is an essential micronutrient for plant growth and development. It is also a crucial element for survival of most of the organisms including humans. Zn acts as cofactor of more than 300 proteins, among which majority are zinc finger proteins, RNA polymerases and DNA polymerases. It is the only metal present in all six enzyme classes (oxidoreductase, transferase, hydrolases, lyases, isomerases and ligases). Being a part of structural or catalytic units, Zn regulates the activities, conformational stabilization and folding of various proteins (enzymes). In addition to this, role of Zn in membrane integrity and stabilization, alleviation of oxidative stress — and as intracellular second messenger has also been reported. Zn is involved in a number of plant physiological processes such as hormone regulation (e.g. tryptophan synthesis, a precursor of IAA), signal transduction via mitogen activated protein kinases, repair processes of PS II complex during photoinhibition and maintenance of CO2 concentration in mesophyll. Peck and McDonald (2010) — confirms the participation of Zn in regulation of Rubisco activity along with alleviation from adverse effects of heat stress in wheat. Thus, Zn essentiality for plant system is illustrated from literature cited above.” [Internal citations removed.]
Zinc activates enzymes that are responsible for the synthesis of certain proteins. It is used in the formation of chlorophyll and some carbohydrates, conversion of starches to sugars and its presence in plant tissue helps the plant to withstand cold temperatures. Zinc is essential in the formation of auxins, which help with growth regulation and stem elongation.
Zinc plays similar roles in animal biochemistry.
Copper: A 2018 paper notes that “copper is an essential element for proper growth and development of plants. Copper in plants is functioning as a catalyst in respiration and photosynthesis. It is vital element for the creation of lignin in plant cell walls and in the case of enzymes responsible for protein synthesis. It also influences the disease resistance and reproduction.” The horticulture site likewise explains the role of copper in plants:
Copper activates some enzymes in plants which are involved in lignin synthesis and it is essential in several enzyme systems. It is also required in the process of photosynthesis, is essential in plant respiration and assists in plant metabolism of carbohydrates and proteins. Copper also serves to intensify flavor and color in vegetables and color in flowers.
Nickel: As for nickel, the same horticulture website explains:
Nickel is a component of some plant enzymes, most notably urease, which metabolizes urea nitrogen into useable ammonia within the plant. Without nickel, toxic levels of urea can accumulate within the tissue forming necrotic legions on the leaf tips. In this case, nickel deficiency causes urea toxicity. Nickel is also used as a catalyst in enzymes used to help legumes fix nitrogen. There is evidence that nickel helps with disease tolerance in plants, although it is still unclear how this happens.
Ideas Have Consequences
I could go on, citing review papers, research papers, biochemistry textbooks, and other authorities explaining how important metals are to the basic functions of numerous proteins. At the very least, it seems an oversight to criticize biology for not using “heavy metals” and yet fail to acknowledge any of the numerous instances where metals are used to fulfill core functions in enzymes and other proteins. To show a full periodic table that highlights only hydrogen, carbon, nitrogen, oxygen, and sulfur, and then to say “Out of all this good stuff, only five chemical elements appear in proteins,” severely underrepresents the degree to which metals are used in biology.
I infer no bad faith or ignorance on Dr. DeBenedictis’s part. Although her Covid-lockdown-era TEDx talk was recorded privately, with opportunities for multiple takes to get things right, I believe what she probably meant was that amino acids don’t use metals, and it was a simple misstatement. But there’s still a lesson here.
Ideas have consequences. When we adopt a materialistic perspective which assumes that biology has no intelligent design, and instead contains fundamental flaws or “gigantic mistakes,” we’ll be quicker to assume that features of biological systems exist for no good reason. This discourages us from investigating why things are the way they are. We’ll be more likely to miss good design features in biology, which is exactly what I think has happened here.
Materialism’s blind spots often exist right where important biological features are found. We’ve certainly seen this blind spot in past thinking about “junk DNA,” where a standard evolutionary framework discouraged research into the functions of this DNA. Today, we know “junk DNA” has key biological functions. But materialistic thinking doesn’t encourage us to understand the design principles that underly biology.
Intelligent design is an idea that has consequences, too. And when it comes to understanding biology, ID is a paradigm that can lead to fruitful research predictions. Those predictions start with the suspicion that there’s probably a good reason why things are the way they are, and task is up to us to figure out what those reasons are.